



INTRODUCTION
Pasture-based automatic milking systems (AMS) require cow traffic (the voluntary movement of cattle around a farm) to enable cows to be milked. In pasture-based systems, this motivation for cow traffic is created by splitting a day’s allocation of feed into multiple distinct allocations of pasture around the farm (Jago et al., 2002). As farms consolidate and become larger in Australia (Dairy Australia, 2012), the distance that cows are walking to access new allocations of pasture is increasing. This presents a unique challenge to AMS as voluntary cow traffic is reduced when the distance that cows are required to walk to access forage increases (Wiktorsson and Sporndly, 2002; Sporndly and Wredle, 2004). Hence, there is renewed interest in utilising alternative forages and the associated capacity to grow greater amounts of forage at a location closer to the milking parlor, particularly when pasture growth rates are low.
The ability to grow 25 t DM/ha/yr in a complementary forage conventional dairy system was highlighted by Farina et al. (2011) exceeding the average Australian dairy farm pasture yield of 7 to 8 t DM/ha (Fulkerson et al., 2005). Their work emphasises the opportunity to incorporate alternative forages into AMS to reduce the average distance of feed from the milking parlor. However, the impact of offering alternative forages on cow traffic in AMS is largely unknown. In a cow preference study (Horadagoda, 2009), cows grazed soybean for a significantly greater time (approximately 70%) than alternative forages such as cowpeas (Vigna unguiculata) and lablab (Lablab purpureus). This work highlighted cow preference for certain forages and the opportunity to utilise the preference for certain forages to modify cow behaviour on AMS farms. Therefore, the aim of this experiment was to investigate the impact on cow traffic of replacing one allocation of pasture with soybean in an AMS system, with the hypothesis that incorporating soybean would increase cow traffic from pasture after it had been depleted and thereby increase milk production through an associated increase in milking frequency.
MATERIAL AND METHODS
All animal experimentation was approved by the NSW Department of Primary Industries Animal Ethics Committee, Menangle, Australia.
Site, animals and treatments
The study was conducted at the AMS research farm, Elizabeth MacArthur Agricultural Institute, Camden (34° 04′S; 150° 69′E) from 28 January 2012 to 11 February 2012. One hundred and eighty mixed age, primiparous and multiparous Holstein-Friesian/Illawarra cows were randomly assigned to two treatment groups (n = 90/group) matched for (means±standard deviation) DIM 174±123 days; parity 2.8±1.9; and milk yield 22.2±8.1 L in a 2 (treatment)×2 (periods) Latin square design.
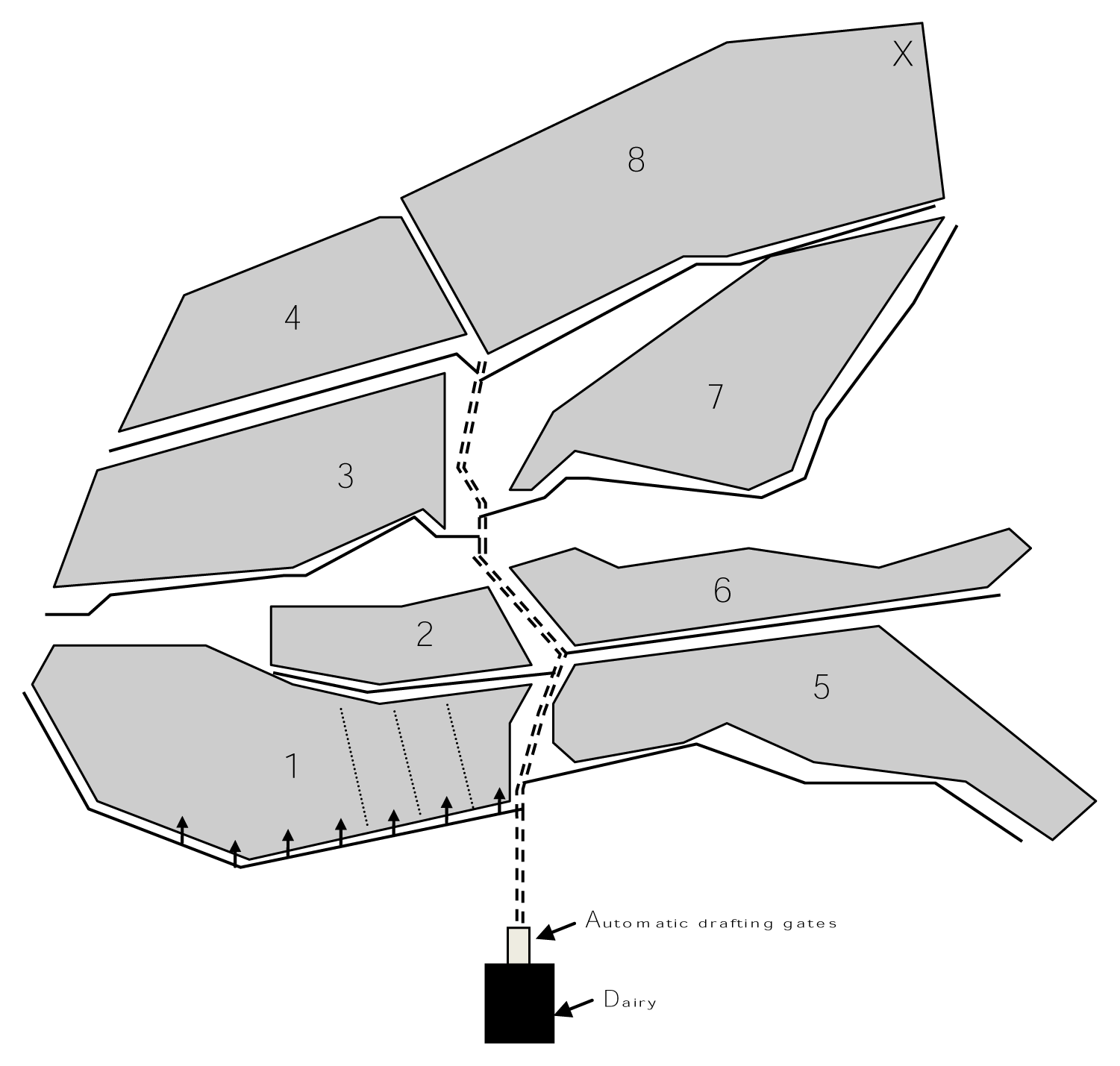
There were two 3 day experimental periods in this study. Both experimental periods followed 5 days of training and adaptation. Each group was allocated to treatments of the legume soybean (Glycine max cv. A6785) or kikuyu grass (Pennisetum clandestinum) (pasture) in period 1 and were offered the alternate treatment in period 2. During the 2 days of training in each period, all cows were batch milked in the morning to ensure that all cows had access to soybean during the day. This training period ensured that all cows were well exposed and familiar with soybean, the laneway layout, location of one-way gates and automatic drafting gates on the AMS farm (Figure 1). For the 3 days of adaptation, cows were split into the two treatment groups.
From 2100 to 0900 h, any cow leaving the milking parlor during adaptation and the experiment was sent to a common ‘night’ pasture allocation (2 kg DM/cow allocated above 1,500 kg DM/ha) (area 1 Figure 1) supplemented with a partial mixed ration of corn (Zea mays) and alfalfa hay (Medicago sativa) (4 kg DM/cow) offered at a feedpad located at the milking parlor (‘Dairy’ Figure 1). From 0900 h until 1500 h, all cows exiting the night allocation or fetched (moved from the paddock to the milking parlor) from the previous afternoon allocation and trafficking to the milking parlor, were either drafted into the milking parlor for milking if the interval since previous milking exceeded 6 h and then to pasture (area 3 Figure 1) or soybean (area 8 Figure 1) at an allowance of 4 kg DM/cow. Cows were drafted directly to the treatment allocation (pasture or soybean) if milking permission was not granted. All cows leaving the milking parlor from 1500 h to 2100 h were drafted to a common (afternoon) pasture allocation (area 6 Figure 1) with an allocation of 2 kg DM/cow. Cows were able to traffic from any paddock to the milking parlor at any time but their destination from the milking parlor was dependent on the time of day (as above). At 1700 h all cows that remained in their assigned treatment paddocks were fetched directly without trafficking through the milking parlor to the common afternoon pasture break. Cows were offered 6 kg DM/cow/d grain based pelleted concentrate after milking with the amount offered at each visit dependant on the number of visitations to the milking parlor.
Pasture and soybean grazing management
Pasture management decision rules were based on matching daily pasture consumption with pasture growth rate (Macdonald and Penno, 1998; Holmes and Roche, 2007) to maintain a whole farm system pasture cover (herbage mass above ground level) of approximately 2,000 kg DM/ha (±200 kg DM), and pre- and post-grazing mass of 2,600 and 1,500 (±200 kg DM), respectively (Garcia and Holmes, 2005). Due to the limited literature on soybean management decision rules, data from the training period were used to determine target post-grazing cover. As 50% of pre-grazing soybean cover was consumed by cows during the training period, cows in the soybean treatment were allocated to this level during the adjustment and experimental periods.
Soybean seed of a germination rate of 92% was inoculated with inoculum H one hour before sowing and then sown at the rate of 80 kg/ha by direct drilling using a Duncan MK3 Renovator on 15th November 2011. Fertilizer (N:P:K:Mo = 5:11:21:7) was applied (50 kg DM/ha) at sowing. The distance between rows was 35 cm to give a target 400,000 plants/ha.
Pasture was located 430 m, and soybean 610 m from the milking parlor.
Measurements and calculations
Forage measurements
Pre- (before cows entered the allocation of pasture) and post- (after the last cow exited the allocation of pasture) compressed pasture height was measured daily by trained staff using a Rising Plate Meter (RPM) (360 mm diameter, 315 g plate weight) fitted with an electronic counter (Farmworks, Palmerston North, New Zealand). Eighty pasture heights were measured in each allocation. These compressed pasture heights were converted to DM yield using the regression equation for kikuyu grass as determined for similar pastures and environment (Garcia et al., 2008). On the day of grazing, pasture samples were hand plucked twice daily at 0800 h and 1500 h while walking across the paddocks to simulate grazing. Soybean pre- and post-grazing cover was determined by harvesting all plants within three 30 cm×30 cm randomly allocated quadrants across each allocation to 5 cm and samples were then stored at 4°C before analysis. The DM difference between pre- and post-grazing mass was determined as an average for the three quadrates and equated to a per hectare basis to determine Soybean consumed. Samples were weighed and then dried at 60°C in a forced-draught oven for 48 h. The dried samples were ground through a 1 mm screen using a Wiley mill and a subsample of pasture and soybean was taken for analysis after thorough mixing.
Pasture and soybean samples were analysed for acid detergent fiber (ADF; van Soest 1963), neutral detergent fiber α-NDF using heat stable alpha-amylase and sodium sulphite (adapted from van Soest et al., 1991), nitrogen (N) concentration by combustion using LECO FP-628 Nitrogen Determinator (LECO Corporation, St. Joseph, Michigan, USA) and water soluble carbohydrate (WSC) content as per Smith (1969). Crude protein (CP) was calculated as N×6.25. Metabolizable energy was estimated using ME (MJ/kg DM) = dry matter digestibility ([DMD, g/100 g]×0.17)−2 (SCA, 1990). Dry matter digestibility of samples was determined by in vitro ruminal incubation (Wang et al., 1999) following estimation of DMD (Clark et al., 1987).
Animal measurements
Cow entry and exit events into and out of paddocks and behavioural activities were determined for each day of the treatment allocation during the experimental period. Eating, walking, rumination, and idling behaviour were monitored every 15-min throughout the day (up to 1500 h) for the first five cows each day that entered in the paddock. The same cows were monitored every 15 min for 1 min per cow to determine bite rate across time. From these data, times for each activity were calculated by multiplying each behaviour activity frequency by a 15-min interval. For the purpose of this experiment, eating behaviour was defined as cows with heads down and engaged in acquiring herbage into the mouth (Gibb, 1998). Milk yield per milking was recorded for each cow and summed to provide daily milk yield.
Statistical analysis
Data were fitted with linear mixed models and parameters, with the exception of grazing behaviour data that was estimated using the restricted maximum likelihood (REML) procedure of GENSTAT for WINDOWS version 14 (Payne et al., 2008).
Day was considered as a random effect as the experimental conditions were assumed to vary randomly from day to day and any cumulative effects were not of interest in this study. Animals were considered as the experimental unit for animal related variables.
The model for animal-related variables with the exception of behaviour data was:
While the model for pasture- and intake-related variables was:
Where Ya and Yb are animal- or pasture- and intake-related variables, respectively.
Standard error of the difference (SED) is reported for comparisons presented.
Logistic regression with random effects was used to analyse behaviour data with numbers of cows grazing, ruminating, idle or walking as the response variable and total cows as the binomial total. Fixed effects included treatment, period and time. Random effects included day. Predicted means were back transformed on the link scale to determine proportions and these were plotted.
RESULTS
During the experimental period, mean rainfall was 4.0 mm/d, maximum air temperature was 27°C, minimum temperature was 16°C and humidity was 76%.
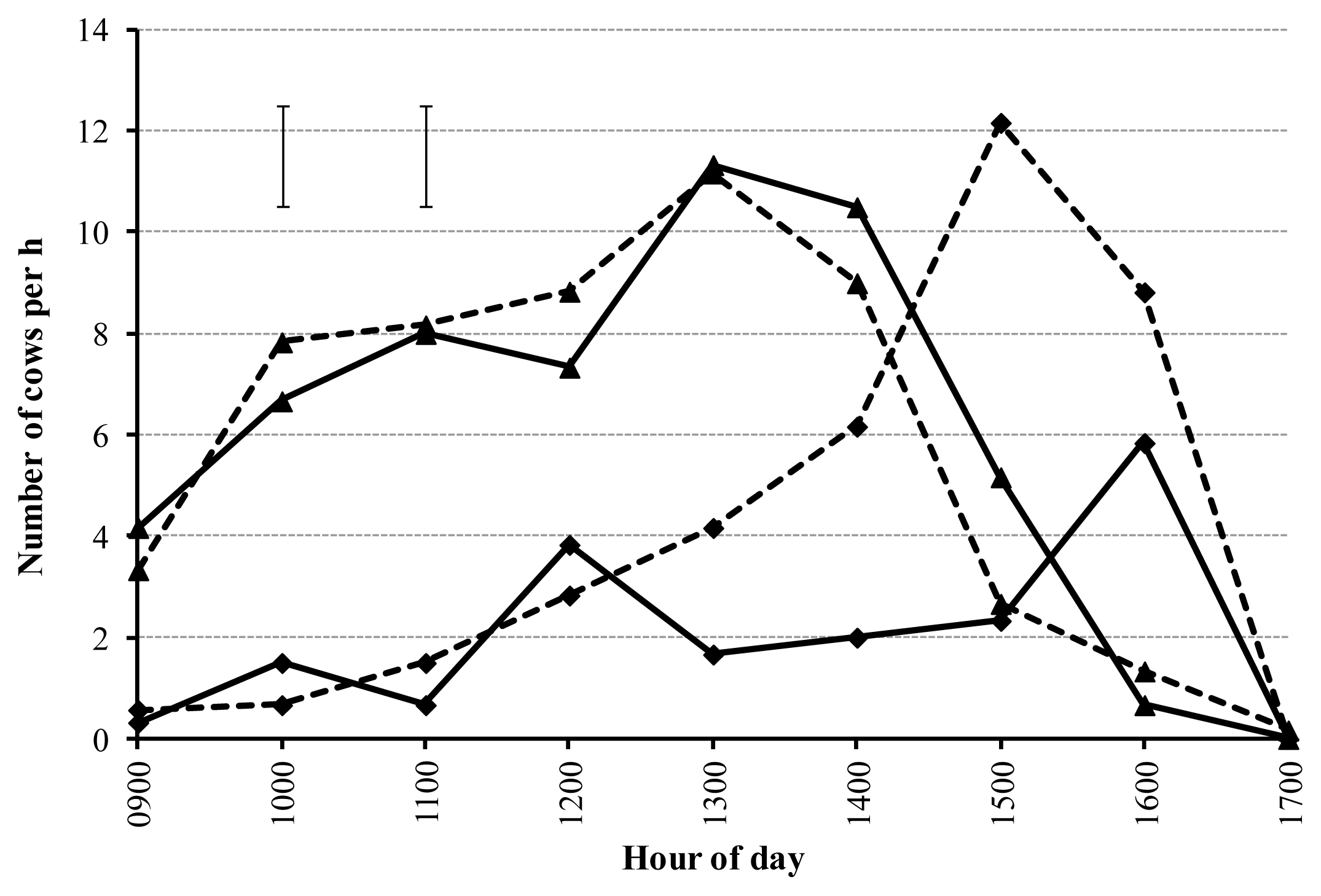
There was no effect (p = 0.83) of forage type on the number of cows entering each respective allocation (mean 53 cows), however, the number of cows entering each allocation varied with time (p<0.01) (Figure 2). The number of cows that exited each forage tended to be greater (p = 0.07) for pasture (34 cows) as compared with soybean (17 cows) and varied with time (p = 0.03). In this regard, cows that arrived at soybean stayed for 25% more time as compared with pasture. Cows entering both treatments reached maximum cow entry at ≈11 cows/h, 4 h after the allocation opened (1300 h). At peak, 12 cows/h exited pasture at 1500 h as compared with the peak of 6 cows/h exiting soybean at 1600 h.
Pre- and post-grazing soybean covers were greater (p< 0.01) than those of pasture (Table 1). Also, the area allocated to soybean was greater (p<0.01) than pasture.
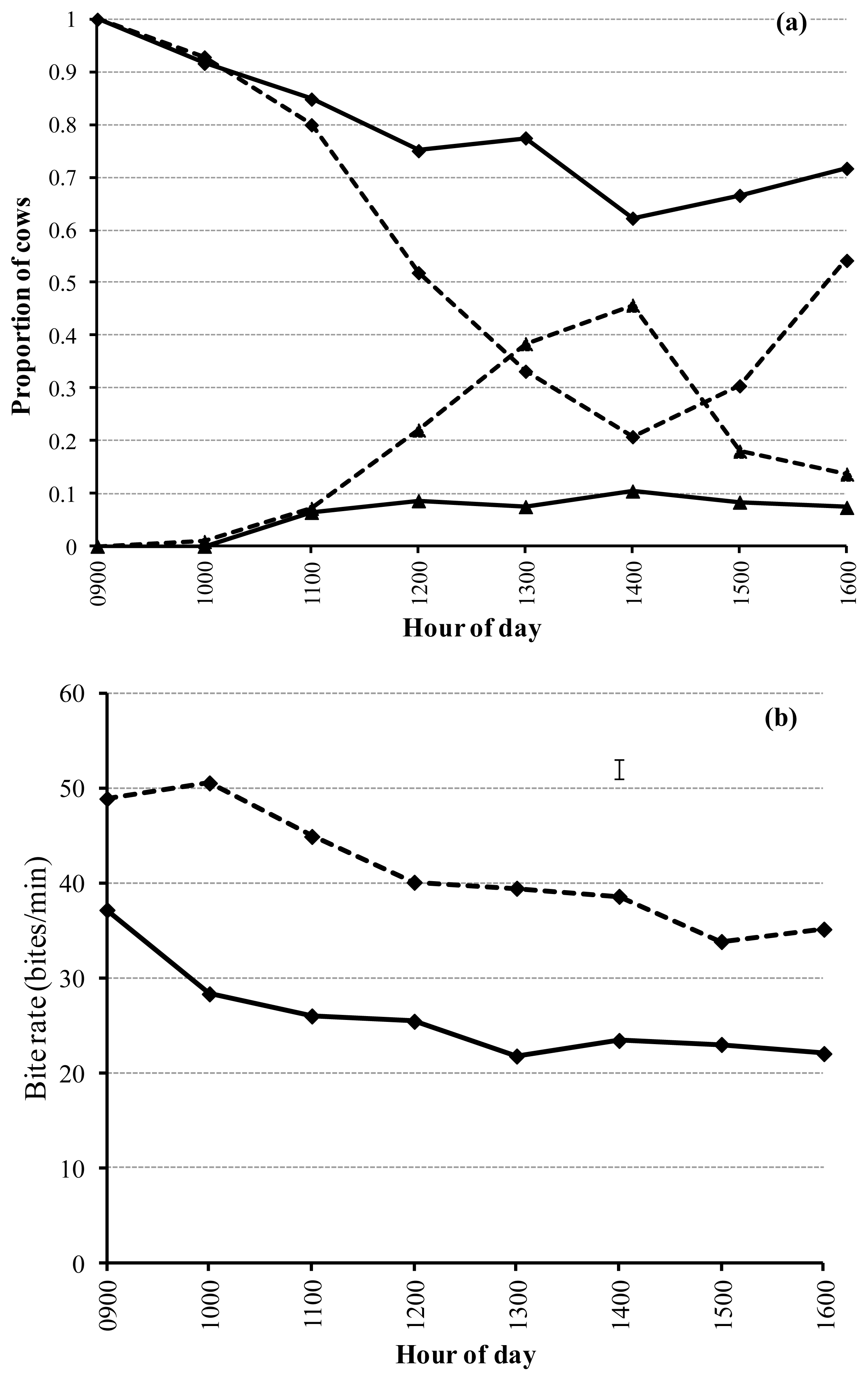
Treatment, time and their interaction were significant (p<0.01) for the proportion of cows within the treatment paddocks that were eating and ruminating (Figure 3a). The greatest proportion of cows eating soybean and pasture occurred when allocations opened (0900 h). By 5 h after allocations opening, the proportion of cows eating in each treatment reached a minimum of 60% and 20% for soybean and pasture, respectively. There were no cows ruminating in the treatment paddocks at the time allocations were opened. The proportion of cows ruminating increased to a maximum of approximately 10% and 45% for soybean and pasture, respectively. Treatment and time were both significant (p<0.01) for the proportion of cows idle (mean 3% soybean and 4% pasture), however, there was no interaction (p = 0.54) between treatment and time. There was no difference between treatments for the proportion of time walking (mean 2%; p = 0.63).
The interaction between treatment and time was significant (p<0.01) for bite rate. Bite rate was greatest for pasture (≈50 bites/min) and soybean (≈40 bites/min) at the time allocations were opened, decreasing to 35 and 22 bites/min for pasture and soybean, respectively, at the end of available grazing time (Figure 3b). Soybean intake (8.5 kg/cow/d) was significantly greater (p<0.01) than pasture (4.7 kg/cow/d). Milk yield per cow was similar (mean ≈18 L/cow/d; p = 0.78) between treatments despite differences in intake.
Pasture DM (23.5%) and NDF (59.9% DM) were greater (p<0.01) than soybean DM (16%) and NDF (34.0% DM). Pasture WSC (8.5% DM) and ME (9.1 MJ/kg DM) were lower (p<0.01) than soybean WSC (10.0% DM) and ME (11.6 MJ/kg DM). There was an interaction (p<0.05) between treatment and period for all other forage tests (Table 2). Generally, the nutritive value of pasture increased from period 1 to period 2 and the quality of soybean decreased.
DISCUSSION
Dairy cows exhibit clear preference for legumes over grass (Torres-Rodriguez et al., 1997; Rutter, 2006) particularly in the mornings (Parsons et al., 1994). Despite this, the number of cows voluntarily moving to soybean and pasture treatments in the current study was similar. Given the preference for soybean over other high nutritive value forages (Horadagoda, 2009), the current work suggests that voluntary cow traffic to allocations of feed will be similar irrespective of forage type. In addition, voluntary cow traffic from allocations of soybean tended to be reduced as compared with pasture, particularly in the late afternoon (1500 h) (Figure 2). The greater distance of soybean from the milking parlor (610-m) as compared with pasture (430-m) may have confounded voluntary cow traffic results in the current work as locating feed at distances greater than 400-m from the milking parlor have been associated with reduced milk production, milking frequency and grazing time (Wiktorsson and Sporndly, 2002). However, more recent data from our research group (Lyons et al., 2013) for cows grazing a predominantly pasture-based diet indicate that this decline occurs at a greater distance of around 800 m. As the treatments within this study were within 800 m from the milking parlor, the divergence in voluntary cow traffic between soybean and pasture back to the milking parlor are more likely caused by differences in nutritive value and animal behaviour between forage types.
It is a common behaviour pattern for grazing cows to eat, drink and then ruminate (Kilgour and Dalton, 1984). However, there was little commonality between soybean and pasture treatments regarding the timing and proportion of cows ruminating and eating in the current study. Cows offered soybean spent a greater proportion of time eating and less time ruminating as compared with pasture (Figure 3a). These differences in behaviour are in line with those expected due to the nutritive value of both species. Leguminous fiber, such as that within soybean, differs from that of pasture in the way that it ferments, breaks down into small particles in the rumen and the greater rate at which it leaves the rumen relative to pasture (Waghorn et al., 1989; Dewhurst et al., 2003). To add to this, it is generally considered that C4 grasses, such as kikuyu, are inferior in nutritive value to C3 grasses (Ulyatt, 1981) as C4 grasses typically have greater concentrations of fiber (Burke et al., 2000). In this regard, NDF in soybean for the current study was approximately half that of pasture (33 vs 60% of DM, respectively; Table 2). Thus, cows offered pasture most likely had a reduced degradability rate in line with the greater rumination time, lower passage rate and therefore reduced intake as compared with soybean.
Greater intakes of soybean (8.5 kg DM/cow/d) as compared with pasture (4.7 kg DM/cow/d) in the current work are supported by the difference in nutritive value between the forages. However, the degree of the response was larger than expected. An 80% increase in DM intake from pasture to soybean found in the current study is well outside the previously reviewed range (−4 to 25%) of DM intake responses for various legumes compared with ryegrass pasture (Waghorn and Clark, 2004). There is limited literature on the additional intake of legumes in comparison with C4 species; however, the additional response for the current work could in part be explained by the much greater nutritive value of perennial ryegrass used in the review as opposed to kikuyu grass (Fulkerson et al., 2007). Also, the work compiled in the review compared the intake of entire legume diets versus perennial ryegrass. As a portion of the total diet in the current study was legume, cows in the soybean treatment had the opportunity to substitute lesser nutritive value partial mixed ration, or kikuyu pasture, offered at other allocations for soybean.
Despite cows spending a greater amount of time on soybean relative to pasture (Table 1), when the average proportion of time grazing as calculated by observation data is related to intake of the treatment herds, the intake rate of both forages was similar (≈50 g DM/min). Thus, the greater intake of soybean was due to the greater proportion of time spent grazing. These findings are in agreement with those of Penning et al. (1997) who showed similar rates of intake for yearling Friesian heifers grazing clover compared to grass, but contrast with the greater intake rates for lactating dairy cattle grazing white clover relative to grass found by others (Rutter et al., 2004; Chapman et al., 2007). Differences in intake rate between legume and grass between studies may be due to differences in the DM% of the forages evaluated. The DM% of soybean (16%) was lower than pasture (24%) in the current study (Table 2) which is the level (16%) at which DM intake is reported to decline (John and Ulyatt, 1987). Thus, despite the greater nutritive value of soybean, the DM intake rate of soybean was likely compromised due to the amount of fresh forage required to be ingested created by the low DM%. To add to the impact of DM%, the morphology of soybean may have further reduced intake rate by reducing bite rate (Figure 3b). Large differences in the approach used by cows to graze soybean and pasture were anecdotally observed in the current study. Cows offered soybean typically selected/grazed the leaves from the stems before grazing down into the stem of the sward. In this regard, cows grazing soybean may have sacrificed bite rate for the ability to select the higher nutritive value leaf portion of the plant.
The high proportion of cows eating soybean throughout the available grazing time, particularly toward the end of the available grazing time, suggests that these cows may have been willing to graze to levels even lower than the post-grazing covers reported here (≈2,400 kg DM/ha; Table 1). This was despite the post-grazing soybean mass being close (51%) to the target residual of 50% depletion (pre-grazing pasture mass ≈4,700 kg DM/ha). As feed is the predominant motivator for cow traffic around AMS farms (Jago et al., 2002), the reduced number of cows voluntarily exiting soybean may have been due to the post-grazing cover target being too high. In this regard, further research is required to determine post-grazing cover targets for soybean, and other alternative forages, to maintain dairy cow intakes whilst optimizing voluntary cow traffic.
The value of legumes for milk production has been attributed to greater intakes by cows and a greater nutritive value as compared with ryegrass (Nuthall et al., 2000; Cosgrove et al., 2001). Waghorn and Clark (2004) reported an increase in milk production of 21% to 48% for cows offered legume relative to cows offered ryegrass. Despite the greater intake of soybean relative to pasture in the current study, there was no effect of forage type on milk production. This unexpected finding may have been due to the substitution of pasture/mixed ration for soybean and/or abrupt changes in rumen environment from the intake of soybean during the day and kikuyu pasture during the night as rumen microflora suited to legume based diets differ from those that digest high levels of cellulose (Waghorn et al., 2007). Thus, greater intakes of soybean in the current study may have been accompanied by reduced in-vivo digestibility of the overall soybean treatment diet whose effect would not be shown using in-vitro DMD (Table 2). In this regard, Woodward et al. (2010) showed a large 24% temporary reduction in milk yield for cows offered lucerne (Medicago sativa L.) when this forage replaced ryegrass. However, after 4 days these cows averaged 20% more milk than those offered ryegrass. Given the large difference in nutritive value between the forages in the current study, the reduction in milk yield as a consequence of a rapid change in the composition of diet may have lasted for longer than 4 days, extending into the experimental period. These findings clearly indicate that further work is required to determine the impact of alternating feed types of differing nutritive value in AMS on feed digestion in the rumen and subsequent animal welfare and milk production.
CONCLUSIONS
Forages that grow a greater amount of DM and that can be grazed with high intake rates are required to help AMS dairy farmers improve voluntary cow traffic, particularly as farm size increases. Despite the previously shown dairy cow preference for legumes, voluntary cow traffic was not increased by offering soybean and intake rates were similar to pasture in the current study. Dairy cows offered soybean had greater intakes as they grazed for a longer proportion of time as compared with pasture, possibly due to the tendency and time required for cows to select out leaf over stem. Also, the milk yield for cows offered pasture and soybean was similar even with the greater soybean intakes. These findings indicate that high nutritive value alternative forages of a more homogenous morphology with less distinction between stem and leaf may be required to increase dairy cow DM intake and intake rate for future AMS farms. Forages such as chicory or forage rape would be potential species to evaluate. Given that there was no increase in milk yield associated with the greater intakes of soybean in the current work, this proposed evaluation should consider the impact of offering forages of differing nutritive value on the rumen environment.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







