The Relationship of Pulmonary Artery Copper Concentrations and Genes Involved in Copper Homeostasis in Cattle, Swine, and Goats*,,**
Article information
Abstract
Liver and pulmonary artery tissue from 5 Angus cross bred steers, 6 goats, and 6 pigs were collected at a commercial abattoir to examine the relationship of pulmonary artery copper (Cu) concentrations and genes involved in copper homeostasis. Liver and pulmonary artery samples were collected at the time of harvest and snap frozen. Liver and pulmonary artery Cu concentrations were determined via flame atomic absorption spectrophotometry and gene expression was determined via real time PCR. Liver Cu concentrations (mg Cu/kg DM±SE) were higher (p<0.01) in cows (396.4±109.1) and goats (181.4±37.0) than in pigs (19.2±3.5). All liver Cu concentrations were within normal ranges and considered adequate for each species. Liver Cu concentration was more variable in cows and goats compared to pig liver Cu concentrations. Pulmonary artery β-hydroxylproline was higher (p<0.01) in cow and pig than goat. Real Time PCR revealed that goat liver atp7a was positively correlated (r2 = 0.92; p<0.01) to liver Cu concentrations while cow and pig atp7a was not correlated to liver Cu concentration. In the pig, liver atp7a concentration was positively correlated to atp7b (r2 = 0.66; p<0.05). Pulmonary artery Cu concentration was highest in cows (14.9±4.7), intermediate in pigs (8.9±3.3), and lowest in goats (3.9±1.1). Goat pulmonary artery Cu concentration was not correlated to ctr1 concentration, however, atp7a concentration was positively correlated with ctr1 (r2 = 0.90; p<0.01). In cow pulmonary artery, loxl1 concentration was positively correlated to eln mRNA concentration (r2 = 0.91; p<0.02). Pulmonary artery CTR1 protein concentration was positively correlated to pulmonary artery Cu (r2 = 0.85; p = 0.03) concentration while negatively correlated to liver Cu (r2 = −0.79; p<0.04). Pulmonary artery Cu concentration was not correlated to concentration of Cu homeostatic genes in the pig. These data indicate that genes involved in Cu homeostasis (ctr1, atp7A, atp7B, loxl1 and eln) are differently regulated in different species.
INTRODUCTION
Pulmonary hypertension in mammals can be caused by oxygen deprivation at high elevations above 1,500 m. Shortage of oxygen due to lower partial oxygen pressure at high altitude induces increased resistance of blood flow in small arteries in the lungs. The right ventricle of the heart compensates for higher blood pressure via a hypertrophic response. The pulmonary artery responds to the increased right ventricle pressure and expands to accommodate the increased output from the right ventricle. At high altitude, bovine and porcine species are known to be more susceptible to pulmonary hypertension than other mammalian species (Rhodes, 2005). It is not clearly understood how certain species can adapt to high altitude while others cannot. Kepler et al. (1971) reported that, in vitro, the amount of smooth muscle present in small pulmonary arteries is positively correlated with pulmonary hypertension and right ventricle hypertrophy in cattle and swine, both of which are highly susceptible to pulmonary hypertension.
Smooth muscle elasticity is maintained by collagen, elastin, glycoprotein, and proteoglycan. Soluble tropoelastins crosslink to form elastin. This reaction is catalyzed by lysyl oxidase (LOX1), a Cu dependent enzyme (Siegel et al., 1970; Harris, 1976). Lesions in the pulmonary artery and aorta, due to Cu deficiency, have been reported in several species (Bennetts and Hall, 1939; O’Dell, 1961; Shields, 1962; Guentner, 1976). These lesions are attributed to a reduction in elasticity of cardiac blood vessels in response to a decrease in LOX1 activity. Furthermore, Cu absorption and homeostatic mechanisms are very different between bovine and porcine species (Wapnir, 1998; Spears, 2003) but both species are sensitive to pulmonary hypertension at high altitude. For this reason, we hypothesized that Cu concentration within the pulmonary artery tissues would relate to Cu homeostatic mechanisms as well as mechanisms involved in elasticity of cardiac blood vessels.
MATERIALS AND METHODS
Animals and tissue collection
All animal tissues were collected from a commercial processing plant in Colorado. Liver, right ventricle, and proximal pulmonary artery samples from mature slaughter weigh cows (n = 5), pigs (n = 6) and goats (n = 6) were collected immediately after slaughter. Post collection, tissues were immediately snap frozen in liquid nitrogen for mRNA and protein isolation.
Copper analysis
Copper concentration was determined via flame atomic absorption. Briefly, tissue samples were dried at 80°C for 24 h. Samples were then dry ashed at 600°C for 12 h. Two milliliters of 3.6 N nitric acid were added to the ashed samples. The crucibles were placed into an Ultrasonic Cleaner (Cole Palmer Model 8845-4) to aid in sample digestion. Samples were then diluted in deionized H2O to fit within a linear range of a standard curve. Liver samples were read at 324.7 nm using a flame atomic absorption spectrophotometer (Varian Model 1275; Walnut Creek, CA). Concentrations of the samples were then determined by a standard curve generated by linear regression using standards of known concentrations.
Beta hydroxyproline analysis
Hydroxyproline was analyzed by colorimetric method described by Switzer (Switzer and Summer, 1971). Briefly, tissue samples were hydrolyzed by 6 N HCl at 110°C for 24 h. Chloramine T was added to hydrolysate and incubated for 20 min. After incubation, sodium thiosulfate was added and mixed. Sample tubes were saturated with potassium chloride. Toluen was added and heated in boiling water for 20 min. Tubes were cooled to room temperature and Ehrlich’s reagents were added. Samples were transferred to 96 well plates and absorbance was measured at 565 nm using plated reader. Hydroxyproline concentration was determined using standard curve.
Total RNA extraction and Real Time PCR
Total RNA from liver and proximal pulmonary artery was extracted using TRI reagent. Total RNA was further purified using an RNeasy kit treated with DNAse I. Single stranded cDNA was synthesized from liver and pulmonary artery total RNA using iScript Reverse transcriptase (BioRad, Hercules, CA) at 25°C/5 min, 42°C/30 min, and then 85°C/5 min. The product was diluted 5× with DNAse/RNAse free water. Diluted cDNA (10 μl) was used as a template for Real Time PCR amplification using SYBR Green (BioRad, Hercules, CA). Glyceraldehyde-3-phosphate dehydrogenase (gapdh) was used as an internal reference for normalization of target genes mRNA concentration. Genes of interest and gapdh primers were designed to generate an amplicon size of 100–200 bp (Table 1). Real Time PCR was performed at 95°C 30 s, 62°C 30 s, and 72°C 15 s for 40 cycles in duplicate using iCycler (IQ5, BioRad, Hercules, CA). Relative gene expression was calculated by the difference in Ct value corrected by housekeeping gene. Following RT-PCR amplification, the PCR products were melted (melting curve) to ensure the quality of amplification. For melting curve analysis, RT-PCR products were incubated for 10 s at each step with an increase in temperature by 0.5°C from 55°C to 95°C in each cycle. PCR products were confirmed by 1% agarose gel and DNA sequencing from Proteomics and Metabolomics Facility at Colorado State University.

Primer sequences for real time PCR
Protein extraction and western blot
Total protein from liver and pulmonary artery was extracted by homogenization with in Laemmli buffer. The homogenate was centrifuged to remove the debris. Sample was mixed with Laemmli buffer and 100 μg protein was loaded on 15% sodium dodecyl sulfate-polyacrylamide gel. Proteins were transferred into nitrocellulose membrane and membrane was blocked in TBS-T (50 mM Tris HCl, pH 7.4, 150 mM NaCl, 0.1% Tween-20) and 5% Non-fat Dried Milk for 1 h. The blot was then incubated with a polyclonal antibody (Gentex rabbit polyclonal CTR1; dilution 1:4,000) for 1 h at room temperature. Following three times of wash with TBS-T, membrane was incubated with secondary antibody (SantaCruz donkey anti-rabbit IgG; dilution 1:10,000) for 1 h. CTR1 protein band was detected by Supersignal chemiluminescence (Pierce Rockford, IL, USA). The intensity of the bands was determined by using densitometry software (UN-SCAN-IT, Silk Scientific Corp.). Antibody in the membrane was then removed by stripping buffer (Pierce, Rockford, IL, USA) and incubated with β-actin (mouse polyclonal, Novus international). CTR1 protein concentration was corrected by β-actin.
Data analysis
Correlation analysis was conducted between the Cu and β-hydroxyproline concentrations of each tissue, and each protein of interest using PROC CORR procedure of SAS.
Messenger RNA concentration levels of genes of interest were calculated by relative gene expression within each group. CTR1 protein concentration was corrected by β-actin. No interaction was observed by treatment and therefore main effect was compared and means were separated using a protected t-test at a significant level of p<0.05.
RESULTS
Copper and β-hydroxyproline
Liver Cu concentration was lowest (p<0.0001) in pig (19.2±3.5 mg of Cu/kg DM) and highest in cow (396.4±109.1 mg of Cu/kg DM; Table 2). Right ventricle Cu concentration, however, was similar across all species and averaged at 16.2±1.3 mg of Cu/kg DM. Interestingly, pulmonary artery Cu concentration showed marginal differences between species (p = 0.0823). Pulmonary artery Cu concentration was highest in cow (14.9±4.7 mg/kg) and lowest in goat (3.9±1.1). Although it was not significant, Cu concentration in the pulmonary artery of the pig was numerically lower than cow. Pulmonary artery Cu concentration was not related to liver Cu concentration in the species studied.

Pulmonary artery β-hydroxyproline concentration followed a similar pattern as pulmonary artery Cu concentrations. Cow and pig β-hydroxyproline (9,708±808 and 10,728±672 mg of β-hydroxyproline/kg DM, respectively) was higher than goat (4,329±364 mg/kg).
Liver
Copper transporter ctr1 mRNA was not correlated with liver Cu concentrations in all three species (Table 3). Atp7A mRNA concentration, however, was positively correlated to liver Cu concentration in goat (r2 = 0.76; p<0.05) while cow and pig atp7a mRNA concentration were not. Atp7b mRNA was positively correlated with atp7a mRNA only in pig (r2 = −0.65; p = 0.06). Elastin mRNA (eln) concentration in the liver was positively correlated to liver Cu concentration in goat (r2 = 0.70; p<0.05), negatively correlated to liver Cu concentration in cow (r2 = −0.88; p<0.05) and was not correlated in the pig.

Correlation coefficients of copper metabolism related genes in the liver of cow, pig and goat
Pulmonary artery
Pulmonary artery Cu concentration did not affect ctr1, atp7a or atp7b mRNA concentration in the pulmonary artery for any specie studied (Table 4). Ctr1 mRNA concentration was positively correlated to atp7a mRNA in pulmonary artery from goat (r2 = 0.91; p<0.05) and cow (r2 = 0.81; p<0.05) but not pig. Atp7b mRNA concentration was not correlated to ctr1 mRNA in cow or pig although there was a trend (p = 0.08) for goats (r2 = 0.5685). Interestingly, ctr1 mRNA showed a negative correlation trend to loxl1 mRNA, which requires Cu as a cofactor (goat: r2 = −0.85; p = 0.08; cow: r2 = −0.4377; p = 0.22) and its target, eln mRNA (goat: r2 = 0.74; p = 0.06; cow: r2 = −0.99; p<0.05) concentration in goat and cow pulmonary artery. Pig eln mRNA concentration was not correlated with ctr1 mRNA concentration in pulmonary artery.

Correlation coefficients of copper metabolism related genes in the pulmonary artery of cow, pig and goat
Cow CTR1 protein concentration in the pulmonary artery was positively correlated (r2 = 0.85; p = 0.03) with pulmonary artery Cu concentration while negatively correlated (r2 = −0.79; p<0.04) with liver Cu (Table 5 and Figure 1). Goat and pig CTR1 protein concentration was not affected by either pulmonary artery or liver Cu concentration although goat CTR1 protein concentration showed similar trend (p = 0.13) with pulmonary artery Cu concentration.

Correlation between CTR1 protein concentration and pulmonary or liver Cu concentration in cow, pig and goat
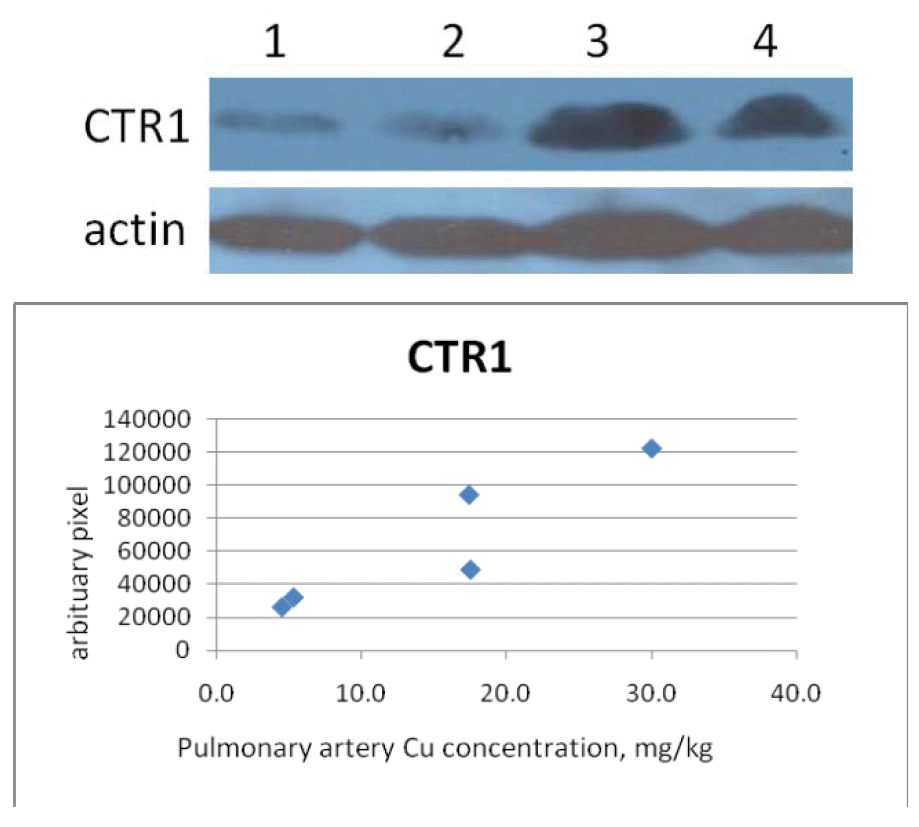
Panel A: CTR1 protein in pulmonary artery from cows at 4.5 and 5.3 mg of Cu/kg DM, respectively (lanes 1 and 2) and 17.4 and 17.5 mg of Cu/kg DM, respectively (lanes 3 and 4) in pulmonary artery. CTR1 protein concentration was corrected by β-actin.
DISCUSSION
Available Cu for absorption in the small intestine varies greatly among species. In general, ruminants have lower Cu absorption from the small intestine (Spears, 2003) relative to non-ruminant animals (Wapnir, 1998). As a consequence, Cu storage in the liver is higher in ruminants than non-ruminant animals to meet the Cu demands by other organs. Cow and goat liver Cu concentration from our study were higher than pig while right ventricle muscle Cu concentration was similar across the species. Copper delivery from liver to the targeted organ is mainly performed by ceruloplasmin in blood (Holmberg and Laurell, 1947; 1948). Absorption of circulating Cu bound to ceruloplasmin into the cell requires Cu transporter (Kuo et al., 2001). CTR1, along with ATP7A, also assists the absorption of copper across the intestinal wall (Pena et al., 1999). After Cu is transported within the cell, Cu trafficking is controlled by several Cu chaperone proteins which includes cytosolic Cu chaperone for superoxide dismutase and Cox 17 and membrane bound ATPase7A and ATPase7B (Prohaska and Gybina, 2004). In our study, right ventricle and pulmonary artery Cu concentration was similar across species regardless of liver Cu concentration. This suggests that Cu delivery mechanism to organs may be regulated to provide similar cellular Cu concentrations regardless of liver Cu concentration across species. While normal liver Cu concentration in swine is lower than ruminants, circulating serum Cu concentrations are approximately twice as high in swine compared to ruminants (Zoubek et al., 1975; Dargatz et al., 1999). This may require different Cu homeostatic mechanisms, i.e. different concentrations of Cu export from the liver as well as different Cu import mechanisms into the cell.
After Cu enters into the cell, Cu is transported by Cu chaperone proteins. ATP7A plays an important role in adding Cu to enzymes that require Cu. One of the enzymes that require Cu is LOXL1 in pulmonary artery. Pulmonary artery, along with aorta, requires the strength as well as elasticity for their normal function. Extracellular matrix elasticity of the artery is formed by network of elastic fibers. Elastin is formed from crosslinks of tropoelastin. Lysyl oxidase is required for crosslink of tropoelastin to from elastin. LOX crosslinks collagen and elastin by deamination of peptidyl lysine residue (Lucero and Kagan, 2006). Lysyl oxidase gene family has five isoforms including LOX and lysyl oxidase like proteins (LOXL) 1–4 (Maki, 2009). So far, LOX and LOXL1 are only known to be responsible for crosslink of collagen and elastin (Borel et al., 2001; Liu et al., 2004). Increased vascular LOX1 activity in hypertensive rats (Ooshima and Midorikawa, 1977) and increased LOXL1 expression in right ventricle of steer with pulmonary hypertension (Han et al., 2008) may suggest that increased activity of collagen crosslinking along with increased deposition of collagen in the pulmonary artery could be responsible for pulmonary hypertension at high elevation.
In our study, ctr1 mRNA was not correlated to liver or pulmonary artery Cu concentration. However, western blot analysis of cow CTR1 in the pulmonary artery showed a positive correlation with pulmonary artery Cu concentration while a negative correlation was observed with liver Cu concentration. No relationship to CTR1 protein concentration was detected in liver or pulmonary artery for goats and pigs (Table 2). These data indicate a possible difference in pulmonary artery Cu metabolism as liver and pulmonary Cu concentrations change. From these data, it appears that as Cu status is decreased in cattle (low liver Cu concentration) that the mechanisms associated with increasing pulmonary artery Cu concentrations increase. Our previous study also demonstrated a negative relationship of ctr1 mRNA concentration in response to liver Cu concentration (Han et al., 2009). It is unclear as to why this occurs.
Loxl1 and eln mRNA was negatively correlated to ctr1 mRNA in goat. Cow and pig pulmonary artery also showed a similar trend. It is not clear if loxl1 mRNA is directly involved in down-regulation of loxl1 and eln mRNA. However, Cu concentration in pulmonary artery from goat was lower than the other species.
Collagen deposition in the pulmonary artery increases with elevated pulmonary arterial pressure (Stenmark et al., 1987; Sun et al., 1994; Tozzi et al., 1994). New born calves kept at 4,300 m for 2 weeks developed pulmonary hypertension with thickened pulmonary artery, mainly in adventitia with increased cellular proliferation of collagen and elastin (Stenmark et al., 1987). Excess collagen in hypertensive pulmonary artery also demonstrated an increase in elasticity (Tozzi et al., 1994) which may contribute to hypertension in pulmonary artery. Although it is not feasible to compare collagen deposition in pulmonary artery across the species, our data shows that collagen content in the cow pulmonary artery was higher in the species most sensitive to pulmonary hypertension at high altitude. This data suggests that the higher concentration of collagen in pulmonary artery could predispose the animal to pulmonary hypertension. It is not clear, however, if collagen deposition in pulmonary artery increases in these species as their pulmonary arterial pressure increases.
ACKNOWLEDGMENTS
We would like to acknowledge Innovative Food (Evans, Colorado) for their generous donation of tissue samples.
Notes
Use of trade names in this publication does not imply endorsement by Colorado State Univ. or criticism of similar products not mentioned.
Mention of a proprietary product does not constitute a guarantee or warranty of the products by Colorado State Univ. or the authors, and does not imply its approval to the exclusion of other products that may also be suitable.