



INTRODUCTION
Meat quality can be defined as a combination of the physical and chemical properties of fresh and processed meat. These properties are represented by the meat color, pH values, water-holding capacity, and cooking loss (van der Wal et al., 1997). Nowadays, antioxidant capacity of meat is also important. Lipid oxidation of meat may lead to decrease eating quality such as odor, flavor, tenderness and juiciness (Fernández et al., 1997). Lipid oxidation is the main non-microbial factor causing the quality deterioration of meat (Fernández et al., 1997; Pradhan et al., 2000; Hernández et al., 2004; Descalzo et al., 2005; Terevinto et al., 2010). Lipid oxidation affects the fatty acids, particularly polyunsaturated fatty acids (Fernández et al., 1997), and the oxidation of unsaturated fatty acids in biomembranes can lead to decreased fluidity of the biomembranes and disruption of the normal membrane structure and function (Dobretsov et al., 1977). So the lipid oxidation damages the integrity of biological membrane which would cut down the water-holding capacity. In addition, oxidation may be linked to meat tenderness (Mercier et al., 2004; Rowe et al., 2004) and might play a role in controlling proteolytic activity of enzymes (Mercier et al., 2004).
The lipid oxidation degradation produces aldehydes, alcohols and other organic compounds that change the meat quality during the storage (Eriksson, 1982). Malondialdehyde (MDA) is one of aldehydes which produced during the lipid oxidation degradation. Also, MDA is thought to be a carcinogenic initiator, and therefore can affect the safety of the food (Fernández et al., 1997). So, MDA contents of meat are very important to lipid oxidation, and the relation between MDA content and lipid oxidation has aroused the interest of investigators (Fernández et al., 1997).
The oxidative stability of muscle is dependent on the balance between the activity of endogenous pro-oxidants (iron, copper, myoglobin, etc.) and antioxidants (antioxidant enzymes, glutathione, etc.) (Gopalakrishnan et al., 1999). Living cells against oxidative processes by several mechanisms of protection, including antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx) (Daun and Åkesson, 2004; Hernández et al., 2004). In addition, SOD and CAT are coupled enzymes (Gatellier et al., 2004; Terevinto et al., 2010). SOD plays an important role in protecting against damage (Chan and Decker, 1994), and decomposes superoxide anion (O2−) to hydrogen peroxide (H2O2) and CAT safely decomposes H2O2 to water (Gatellier et al., 2004; Mercier et al., 2004). GPx is considered the major peroxide-removing enzyme located in the cytosol (Chan and Decker, 1994; Decker and Xu, 1998). And GPx can decompose both H2O2 and lipoperoxides formed during lipid oxidation (Gatellier et al., 2004).
Laiwu pig is one of Chinese indigenous black pig breeds and is mainly reared in Shandong province. Previous research showed that SOD activity of Longissimus dorsi muscle in Laiwu pigs was significantly higher than that in Large White pig (Li et al., 2010). Otherwise, the GPx4 activity of Longissimus dorsi muscle in Laiwu pigs also was significantly higher than that in Large White pig (Chen et al., 2011). So, the purposes of this study were: i) to quantify and compare the major antioxidant enzymes (SOD, CAT and GPx) activities, and ii) to quantify and compare the lipid oxidation by measuring the MDA contents in porcine Longissimus dorsi muscle when stored over a period of six days (d 1 to d 6) at two different temperatures (4°C and 20°C).
MATERIALS AND METHODS
Animals and samples preparation
Twelve pigs (Laiwu pig, n = 6; Large White pig, n = 6) were used in this experiment. All pigs were reared at Laiwu pig breeding farm at Laiwu city and were fed a commercial pig feed and water ad libitum. The pigs were handled according to the ‘Regulations on Administration of Hog Slaughter’ and ‘Good manufacturing practice for pig slaughter (GB/T 19479-2004)’. All pigs were slaughtered at a local commercial abattoir (LIURUN SHIPIN Co., Ltd.) about 6 months of age following standard industrial procedures and the average carcass weight of the pigs was 80±5 kg. The Longissimus dorsi muscles (whole lions of the right carcasses) were obtained from carcasses, and were sliced into 2-cm thickness chops perpendicular to the fiber direction. The chops were stored at two different temperatures (4°C and 20°C) for 6 days (1 to 6 d), and the samples stored (20°C) in the dark. The activities of SOD, CAT and GPX, and MDA contents of the samples of each day were measured during storage.
Assays of antioxidant enzyme activities
A 0.4-g muscle sample was homogenized in a tissue homogenizer (ULTRA-TURRAX® T18 basic, IKA, Germany) with 3.6 ml of 0.86% NaCl for 3×10 s at 23,000 rpm in an ice bath and the homogenate centrifuged at 4°C for 15 min at 7,000×g. The supernatant was filtered through glass wool and used to determine antioxidant enzyme activities. Superoxide dismutase, catalase and glutathione peroxidase assay kits (Nanjing Jiancheng Bioengineering Institute, Jiangsu Province, China, http://www.njjcbio.com/) were used for the assay of SOD, CAT and GPx activities according to the manufacturer’s instructions. One unit of SOD activity is defined as the amount of enzyme in each milliliter of the reaction solution at 50% SOD inhibition at 37°C. One unit of CAT activity is defined as the amount of enzyme in one milligram tissue protein consumption of 1 μmol H2O2 per second at 37°C. One unit of GPx activity is defined as the net amount of the enzyme capable of hydrolyzing 1 μmol of GSH (Glutathione) per min at 37°C.
Assays of MDA contents
The MDA assays kits (Nanjing Jiancheng Bioengineering Institute, Jiangsu Province, China, http://www.njjcbio.com/) were used to assay the MDA contents of Longissimus dorsi muscle of Laiwu pig and Large White pig. MDA can react with thiobarbituric acid (TBA), and obtaining a red pigment which have the maximum absorbance at 532 nm. The MDA values were expressed as nmol/mg protein.
Statistical analysis
All values are reported as mean±standard error of mean (SEM) for three measurements on each muscle sample. The data were analyzed by the procedure Repeated Measures Analysis of Variance of Statistical Analysis Systems statistical software package (Version 9.2; SAS Inst., Inc., Cary, NC, USA). The correlations between the antioxidative enzymes activity and MDA content were characterized by the PROC CORR procedure of the SAS software. The p-value <0.05 was considered to be statistically significant.
RESULTS
Changes in SOD activities during postmortem storage
Data on the SOD activities of porcine muscles are presented in Table 1. From the table, it shows the storage temperatures significantly affected the SOD activity and the SOD activities of porcine muscles were decreased during storage days. The SOD activities of porcine Longissimus dorsi stored at 4°C were significantly (p<0.01) higher than that stored at 20°C (Table 1). The SOD activity of porcine muscle stored at 4°C decreased slowly when stored a period of 1 to 6 days. However, compared with stored at 4°C, the decrease trend of SOD activity stored at 20°C was faster.
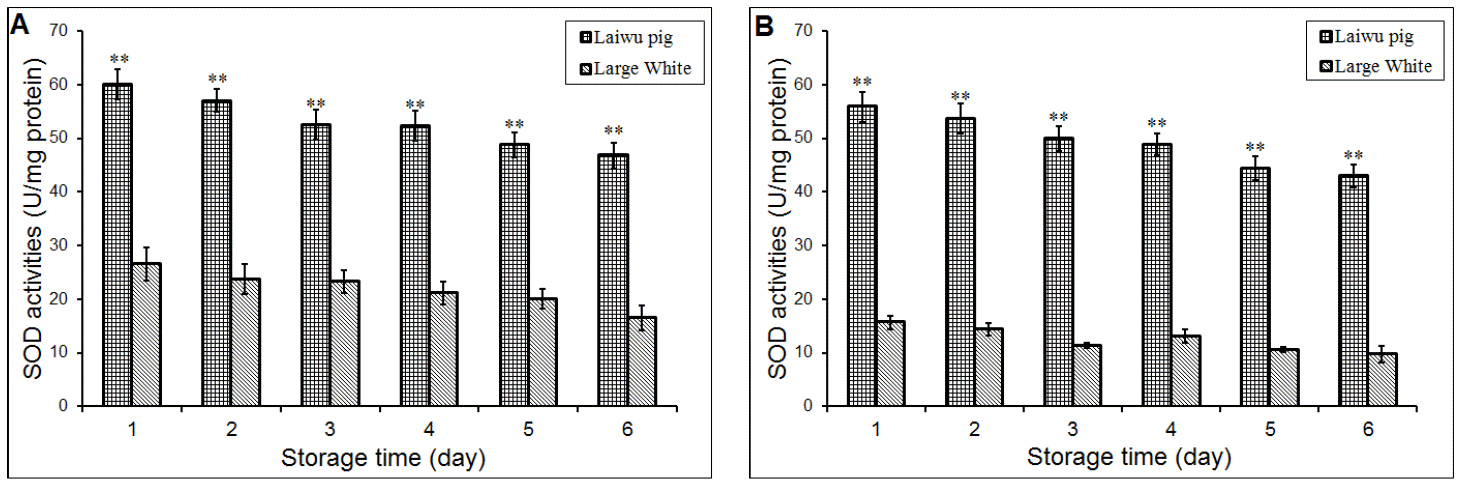
Furthermore, pig breeds also can affect the SOD activity; and Figure 1 shows the effects of pig breeds on the changes of SOD activities of muscle stored at 4°C (Figure 1A) and 20°C (Figure 1B). As shown in Figure 1, the activity of SOD of Laiwu pigs was significantly (p<0.01) higher than Large White’s SOD activity when the muscles samples stored at two temperatures (Figure 1). Compared with Large White pig, Laiwu pig as a Chinese indigenous pig breed showed strongly SOD activity, and it suggested that Laiwu pig can more effectively inhibit lipid oxidation and have a long shelf-life after slaughtered.
Changes in CAT activities during postmortem storage
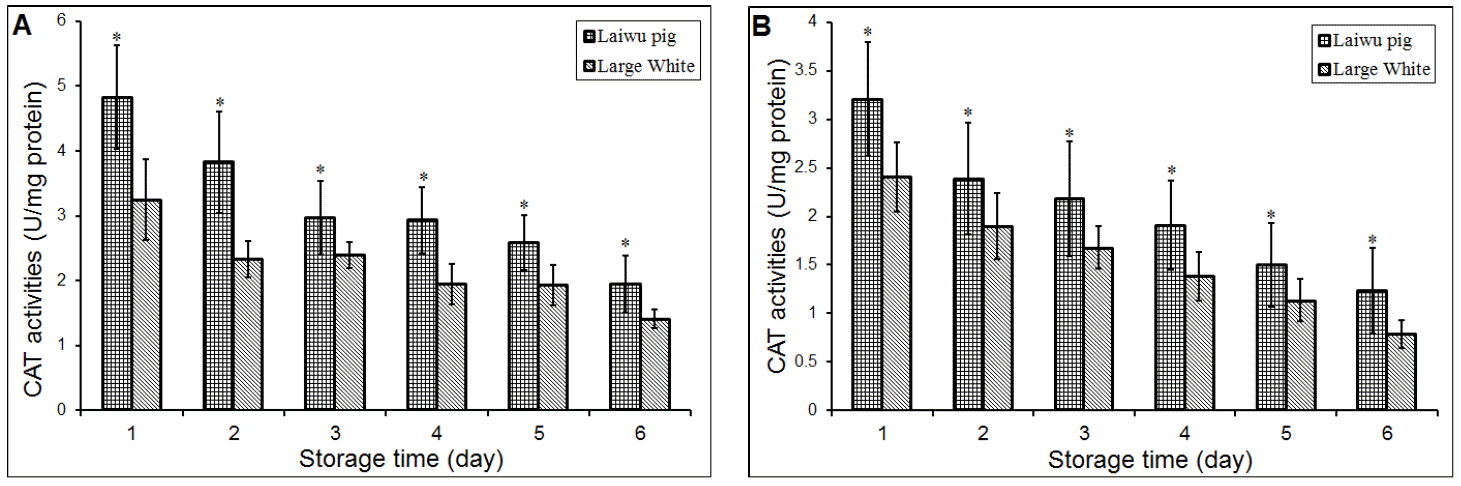
The changes in CAT activity were deceased drastically during storage and the data are presented in Table 2. Storage temperature and time significantly affected the CAT activity. The CAT activity of porcine Longissimus dorsi stored at 4°C was significantly (p<0.01) higher than that stored at 20°C and there was significantly changed in CAT activity of porcine Longissimus dorsi at different storage days. On the other hand, pig breed also can affect the CAT activity (Figure 2). Laiwu pig as a Chinese native breed shows higher CAT activity when the porcine muscles stored at 4°C and 20°C. Muscle CAT activity was significantly (p<0.05) higher in Laiwu pig than in Large White pig when the muscles stored at 4°C and 20°C (Figure 2). Like the Laiwu pig, the CAT activity of Large White pig was also higher stored at 4°C than that stored at 20°C.
Changes in GPx activities during postmortem storage
Table 3 shows the GPx activity when the muscles stored at different time. The activity of GPx enzyme decreased drastically during storage. The storage days significantly affected the GPx activity from Table 3. And the muscle GPx activities stored at 4°C were significantly (p<0.01) higher than that stored at 20°C in the two pig breeds.
In addition, effects of pig breed on the changes in GPx activities of Longissimus dorsi muscle during storage were showed in Figure 3. In Laiwu pigs, the GPx activities decreased drastically when the muscles stored at 20°C. Compared with stored at 20°C, however, the GPx activities in Laiwu pigs were only slightly affected during storage at 4°C. Furthermore, muscle GPx activity in Laiwu pig was significantly (p<0.01) higher than that in Large White pig at the two storage temperatures (4°C and 20°C).
Comparative analysis in muscle MDA contents
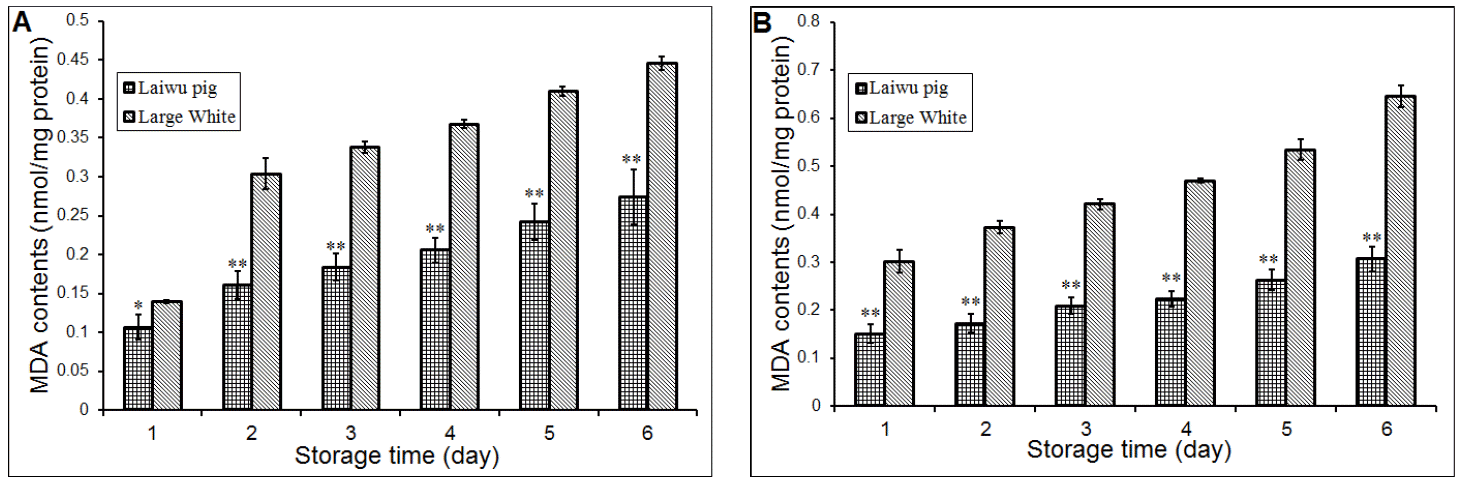
Oxidation in meat usually leads to deterioration in quality. Lipid oxidation values were measured by MDA content in this experiment. During meat maturation for 6 days, there was an increase in MDA content and data on the MDA contents of the muscles are presented in Table 4. As shown in Table 4, the MDA contents were increased drastically during storage. The MDA contents were significantly affected by storage times; and the MDA contents stored at 4°C were significantly (p<0.01) lower than that stored at 20°C. Also, pig breed is one important factor on the MDA content of porcine Longissimus dorsi muscle during storage. The MDA contents of Large White pig were significantly (p<0.01) higher than Laiwu pigs when the muscles stored at 4°C and 20°C (Figure 4).
Relationship between antioxidant enzymes and MDA contents
Measurement of the antioxidant enzyme activity showed a better protection to the lipid oxidation. So, we analyzed the correlation between antioxidant enzyme activity and MDA content (Table 5). The results showed negative correlations were obtained between antioxidant enzyme activities and MDA contents, especially the correlation between SOD activity and MDA content during storage time (r = −0.75; p<0.0001).
Antioxidant defenses in meat are composed by non-enzymatic system like vitamin E and enzymatic system like SOD, CAT and GPx (Descalzo and Sancho, 2008). After the death of pigs, the enzymatic system probably acts to prevent the lipid oxidation in the meat maturation. Our results show the MDA content increase with increasing storage time, and antioxidant enzymes play an important role in protecting lipid oxidation.
DISCUSSION
Oxidation rate of postmortem muscle depends on the anti-oxidative capacity in the animal; and also, oxidation of muscle components postmortem can be retarded by the action of endogenous antioxidative enzymes, especially SOD, CAT and GPx, which seem to be relatively stable during refrigerated storage. Also Renerre et al. (1996) found that SOD activity in Psoas major was higher than that in Longissimus lumborum. These results indicated that there is a natural protection which decreases the deleterious effects of oxy radicals. The reported results were in accordance with previously published data on these varieties. In our results, during 6 days storage at 20°C, muscle SOD activity of Large White pig decreased significantly (p<0.01) between d 1 (15.68 U/mg protein) and d 6 (9.71 U/mg protein) postmortem. Similar to this result, Laiwu pig’s muscle SOD activity decreased significantly (p<0.01) from 55.89 U/mg protein to 42.99 U/mg protein during 6 days storage (Figure 1).
Previous research reported that exogenously added CAT to beef samples failed to prevent lipid oxidation (Pradhan et al., 2000). Descalzo et al., (2000) found CAT activity in Psoas major muscle in beef still remained stable after 9 days storage. And, Renerre et al. (1996) also found CAT activity in Diaphragma muscle didn’t change significantly after 8 days refrigerated storage, but it was higher (p<0.05) than that in another three muscles (Psoas major, Longissimus lumborum and Tensor fasciae latae). In our results, however, CAT activities significantly changes (p<0.05) during storage at two different temperatures; Laiwu pig as a Chinese indigenous pig breed, and its muscles shows higher CAT activity which was significantly higher than Large White pig during storage at 4°C and 20°C (Figure 2).
GPx activity in meat samples is controversial (Descalzo and Sancho, 2008). The GPx activity was strongly affected by the diet in Charolais cattle (Gatellier et al., 2004), and also influenced by the degree of unsaturation of the diet in chicken (Maraschiello et al., 1999). However, in our results, there was significantly (p<0.01) decrease in GPx activity in porcine Longissimus dorsi muscle in Laiwu pigs and Large White pigs. The muscle GPx activity was affected by pig breeds and storage temperatures. Muscle GPx activity in Laiwu pigs was significantly (p<0.01) higher than that in Large White pig at the same storage temperature. On the other hand, muscle GPx activity of Laiwu pigs decreased significantly (p<0.01) between d 1 (8.77 U/mg protein) and d 6 (3.64 U/mg protein) postmortem stored at 20°C, also Large White’s GPx activity between d 1 (4.45 U/mg protein) and d 6 (0.84 U/mg protein). There are the same results when the muscles stored at 4°C (Figure 3).
MDA is the principal and most studied product of polyunsaturated fatty acids per-oxidation. The measurement of MDA is important for the assessment of oxidative stress as MDA has been used as a biomarker of lipid oxidation for more than 30 years (Del Rio et al., 2005). In the present research, the muscle MDA contents were significantly (p<0.01) affected by pig breed and storage temperature. The Laiwu pig’s muscle MDA contents were lower than large white’s; and also, muscle MDA contents stored at 4°C were lower than that stored at 20°C (Figure 4).
Recently, Xia et al. (2009) reported there was a significantly (p<0.05) increase in TBARS when the porcine Longissimus dorsi muscle subjected to multiple freeze-thaw cycles, and this indicated an increase in the lipid oxidation. On the other hand, the apparent relationship between protein carbonyls and TBARS indicated a strong likelihood that some dicarbonyl compounds derived from lipid oxidation, notably MDA, formed complexes with proteins (Li and King, 1996; Xia et al., 2009). In this work, we found that there was significantly (p<0.01) increase of the muscle MDA contents during storage, and it indicated an increase in the lipid oxidation in porcine Longissimus dorsi muscle (Figure 4). In addition, the development of lipid oxidation may vary according to the age, muscle type, breed and feeding regime (Descalzo and Sancho, 2008). The lipid oxidation produces MDA; and the Longissimus dorsi of Laiwu pig breed shows lower MDA content than Large White. So, the lower MDA content of Laiwu pig reflects there was lower level of lipid oxidation in our experiment. Anyway, changes in MDA content could have occurred during storage at the two different temperatures (4°C and 20°C).
SOD, CAT and GPx are very important antioxidative enzymes which can prevent oxidation reactions to improve meat quality and extend the shelf-life. In our results, there will be a strongly antioxidative capacity, if there are higher enzyme’s activities of SOD, CAT and GPx, and lower MDA contents. So, improving the antioxidative enzyme’s activity of muscle by selective breeding is very important to the meat quality and safety.
CONCLUSIONS
In summary, this study evaluated the changes in the main antioxidative enzymes activity and MDA contents in pork meat during storage. These results indicate that the enzyme’s activity of SOD, CAT and GPx was decrease during the storage at two different temperatures, and the MDA contents were increase during the storage. There were the higher antioxidative enzymes activities and lower MDA contents when the pork meat stored at 4°C. It suggested 4°C is more suitable for meat storage compared with 20°C. However, the antioxidative capacity of the Chinese native pig, Laiwu pig, is stronger than Large White’s at all the storage time. In addition, antioxidant enzymes activities were negative correlated with the MDA contents. These results indicate pork muscle samples with higher antioxidant enzymes activity would be less susceptible to lipid oxidation.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







