




INTRODUCTION
The gastrointestinal tract mucosa constantly confronts a variety of potentially injurious agents involving pathogenic bacteria, normal bacterial floras, and even toxic dietary factors, which usually cause infection and inflammation and have been implicated in essential barrier function failure including inflammatory bowel disease (Irvine and Marshall, 2000). Nutritional modulation offers an appropriate approach to prevent mucosal barrier injury.
Herbal ingredients are deemed significant nutrients to improve animal health in China. Acanthopanax senticosus (AS) is a well-known shrub native to far eastern areas of Russia and the northern regions of Japan, Korea, and China (Deyama et al., 2001). Polysaccharides (ASPS) are a major active ingredient of the aqueous extract isolated from AS. Our previous research revealed that the anti-inflammatory role was closely linked to down-regulatory expression of pro-inflammatory cytokines (Han et al., 2014). Due to inflammatory intestinal condition resulting in compromised intestinal barrier function preliminary research was conducted to determine the effect of ASPS on intestinal function. From this, we hypothesized that ASPS would sustain positive changes during intestinal mucosal damage during inflammation conditions.
In the present study, we employed a well-documented intestinal injury model in the mouse by injecting Escherichia coli lipopolysaccharide (LPS) to assess the effect of ASPS on intestinal morphology, gut digestive profile, intestinal tight junction proteins, and mucous layer secretions that characterize intestinal mucosal barrier function, and to probe into the potential mechanisms by which ASPS protects the intestinal mucosal against injurious agents.
MATERIALS AND METHODS
Acanthopanax senticosus polysaccharide preparation
ASPS were prepared from the root of Acanthopanax senticosus using Ethanol precipitation method as previously described (Han et al., 2014). Protein was removed by Sevag method (Staub, 1965) and the polysaccharide content at post-purification was 92.7% as determined by the phenol sulfuric acid method (DuBois et al., 1956). Monosaccharide composition analysis of ASPS by ion chromatography according to the method of Ou et al. (2006) showed that it was a type of heteropolysaccharide composed of glucose, galactose, arabinose, mannose, rhamnose, and xylopyranose.
Mice care and experimental design
All studies were approved by and performed in compliance with the guidelines of the Animal Care and Use Committee of Liaoning Province, China. Male Kunming mice with SPF grade at 5 wk of age (Changsheng Life Sciences Co. Ltd., Changchun Jilin, China) were kept under stable temperature (21┬░C┬▒1┬░C) and humidity (45%┬▒2%) with a 12-h-light/dark cycle and free access to food and water. Following the adaption to the housing condition for 1 wk, mice were distributed into random groups of 6 mice (3 mice/cage) and treated as follows: mice were given ASPS aqueous solution at 300 mg/kg body weight (BW) by daily oral gavage for continuous 14 days (ASPS+LPS group), the others were given an equivalent amount of sterile saline (Control group) and (LPS group). On the morning of 15 days, mice in LPS group and ASPS+LPS group were injected intraperitoneally with Escherichia coli LPS (E. coli Serotype 055: B5, Sigma, Saint Louis, MO, USA) at 4 mg/kg BW, and others in Control group were given an equivalent amount of sterile saline. ASPS dose was determined in accordance with optimal dose obtained in piglets of our previous study (Han et al., 2014), by using dose conversion formula for different animals in pharmacological studies (Xu et al., 2002).
Intestinal sample collections
Following 4 h post-injection with LPS or saline, all the mice were killed by cervical dislocation. Intestinal segments measuring 1 cm and 10 cm in length were excised respectively from the proximal jejunum in each mouse. The 1-cm intestinal segments were washed with ice-cold phosphate-buffered saline (PBS) and the liquid removed with filter paper, and then fixed in 10% formaldehyde for histology sections to observe intestinal morphology. The 10-cm intestinal segments were opened longitudinally and flushed with ice-cold PBS and scraped to gather mucosa samples, which were rapidly freeze-clamped in liquid nitrogen and then stored at ŌĆō80┬░C.
Intestinal morphology
Tissue slices were prepared by hematoxylin and eosin straining according to the method of Luna (1968). In brief, intestinal segments were processed by being fixed, dehydrated, paraffined, embedded, sliced and stained. Five observations were made on each slice and in each observation two villi that were well-stretched and parallel were measured quantitatively for analysis of villus height (the vertical distance from intestinal gland to the villus tip), crypt depth (the vertical distance from the opening of intestinal gland to the bottom), and villus height:crypt depth ratio (VCR).
Intestinal mucosa cells count
Intraepithelial goblet cells and lymphocytes were assayed by hematoxylin and eosin straining. Tissue slices were prepared according to the method of Luna (1968). In brief, intestinal segments were processed by fixed, dehydrated, paraffined, embedded, sliced and stained. Lymphocytes near the side of the intestinal epithelial cells (IEC) base membrane were characterized as small cells with hyperchromatic large nucleus. Goblet cells located between IEC were characterized by relative large cell with hyperchromatic small nucleus. These two kinds of intestinal mucosa cells were observed among one hundred IEC of the third pair of well-stretched and parallel villi in each of the five observation fields of a Axio Scope A1 of ZEISS biomicroscope (Oberkochen, German)
Determination of biochemical parameters of intestinal mucosa
Mucosal disaccharidase activity including lactase, sucrase, and maltase were measured by glucose oxidase-peroxidase conjugated method according to Hou et al. (2006) using commercial kit (Nanjing Jiancheng Biological Product, Nanjing, China), and the values of enzyme activity are expressed as U/mg protein. The activity of diamine oxidase (DAO), as well as the concentration of tumor necrosis factor (TNF)-╬▒ and intestinal trefoil peptide (ITP) were analyzed using enzyme-linked immunosorbent assay with commercial kits (R&D Inc., Los Angeles, CA, USA).
Real-time polymerase chain reaction detection
In brief, total RNA was extracted from intestinal mucosa using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturerŌĆÖs instructions. The quantity and quality of the RNA were verified by spectrophotometry (UV-300; Applied Photophysics, Manchester, England) and agarose electrophoresis. cDNA synthesis was performed using a Promega GoScript reverse transcription system (Promega, Madison, WI, USA) according to the manufacturerŌĆÖs instructions. Gene-specific primers for ╬▓-actin (forward: 5ŌĆ▓-TCCCTGTATGCCTCTGGT-3ŌĆ▓; reverse: 5ŌĆ▓-TGTCACGCACGATTTCC-3ŌĆ▓; GenBank No: NM_007393), heat shock protein (HSP70) (forward: 5ŌĆ▓-CACAAGAAGG ACATCAGCCAGAA-3ŌĆ▓; reverse: 5ŌĆ▓-GGAACAGGTCCG AGCACAGC-3ŌĆ▓; GenBank No: NM_010478); epidermal growth factor (EGF) (forward: 5ŌĆ▓-CCCTCGTCA CTGGTTGT -3ŌĆ▓; reverse: 5ŌĆ▓- TTCTGTCCCATCATCGTC -3ŌĆ▓; GenBank No: NM_010113); epidermal growth factor receptor (EGFR) (forward: 5ŌĆ▓- AATCCCTTTGGAGAA CCT -3ŌĆ▓; reverse: 5ŌĆ▓- TGAATCGCACAGCACC -3ŌĆ▓; GenBank No: NM_007912) were synthesized by Dingguo Changsheng Biotech Co. (Beijing, China). The polymerase chain reaction (PCR) quantitative analysis was carried out on PCR system (T300, Biometra, Jena, Germany) and performed in a 20 uL reaction system containing 7 ╬╝L nuclease-free water, 0.4 ╬╝L up-premier (10 ╬╝mol/L) and 0.4╬╝L down-premier (10 ╬╝mol/L), 10.0 ╬╝L qPCR master mix (2├Ś). The PCR cycling conditions were a pre-denaturation cycle at 95┬░C for 10 min firstly, followed by 40 cycles at 95┬░C for 15 s and 60┬░C for 60 s, then 1 cycle at 95┬░C for 15 s, 60┬░C for 60 min and 95┬░C for 15 s. The relative expression of target genes relative to ╬▓-actin was calculated by using the 2ŌłÆ╬ö╬öCT method (Livak and Schmittgen, 2001).
Western blot for protein expression analysis
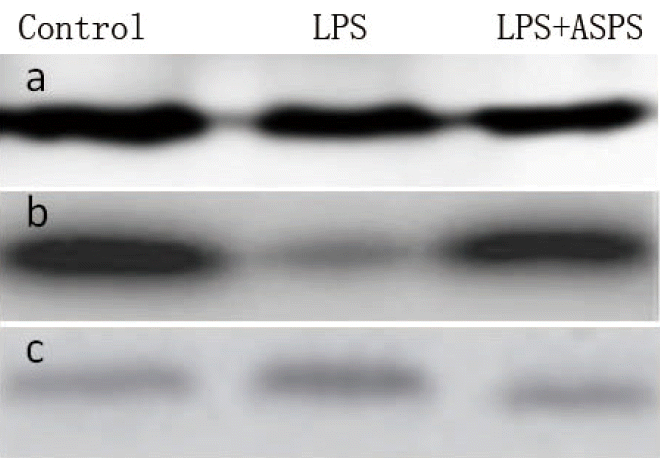
In brief, intestinal mucosal samples were homogenized in 1 mL lysis buffer and centrifuged at 12,000├Śg at 4┬░C for 30 min for supernatants collection, and then the protein concentrations were measured using phenol reagent method. An equal amount of proteins samples (20 to 40 ╬╝g) was separated on a 10% or 12% reducing polyacrylamide gel and transferred onto polyvinylidene difluoride membranes, and incubated with primary antibodies. Immunoblots were blocked with 3% bull serum albumin (BSA) in Tris-buffered saline for 70 min at room temperature and incubated overnight at 4┬░C with the specific primary antibodies in Tris-buffered saline and 0.05% Tween 20 containing 1% BSA. The specific primary antibody included rabbit anti-claudin-1 (1:1,000; Proteintech, Chicago, IL, USA) and anti-occludin-1 (1:1,000; Proteintech, USA). Blots were washed with the same buffer and then incubated with anti-rabbit horseradish peroxidase-conjugated secondary antibody (1:5,000; Proteintech, USA) for 120 min at room temperature. Membranes were washed in the same buffer and antigen-antibody complexes visualized by using an ECL kit (Pierce, Waltham, MA, USA). The density of the bands was quantified using the Image J 1.46r analysis software.
Statistical analyses
Individual mouse was used as experimental unit for analysis of experimental indices. Data were analyzed by one-way analysis of variance. Differences between the groups were analyzed by Duncan multiple comparisons Tests. The results were expressed as means┬▒standard error of mean (SEM), and a 5% level of probability was considered significant for all analyses. The statistical calculations were carried out by using SPSS 19.0 software (Cabit Co., LTD, Shanghai, China).
RESULTS
Intestinal morphology and mucosal cell count
The mice in LPS group had significantly lowered villus height (24.55% lower, p<0.05) and VCR (38.78% lower, p<0.05), as well as greater crypt depth (21.69% greater, p<0.05) in jejunum compared with the mice in control group (Figure 1; Table 1). Correspondingly, ASPS+LPS group significantly increased the villus height (20.84% higher, p<0.05) and VCR (42% higher, p<0.05), as well as decreased the crypt depth (15.55% lower, p<0.05) compared with the LPS group. Intestinal goblet cells number in LPS group was not significantly different from the mice in control group (Figure 2). The mice in ASPS+LPS group showed significantly increased goblet cells (24.89% higher, p<0.05) relative to those in LPS group. Intraepithelial lymphocytes were not significant different among different groups (p>0.05).
Proteins expression of intestinal tight junction
Mucosal Occludin-1 proteins expression was significantly lower (53.33% lower, p<0.05) in LPS group compared with control group although intestinal claudin-1 protein expression did not differ between the two groups (p>0.05) (Figure 3; Table 2). Correspondingly, the mice in ASPS+LPS group demonstrated significant increase in intestinal occludin-1 (71.43%, p<0.05) proteins expression compared with those in LPS group. ASPS pre-treatment had no effect on intestinal claudin-1 protein expression.
Mucosal disaccharidase activity and other biochemical parameters
The mice in LPS group had significantly lower lactase (20% lower, p<0.05) and sucrase (50% lower, p<0.05) activities compared to the mice in control group (Table 3). Compared to the LPS mice without ASPS treatment, the mice in ASPS+LPS group had significant higher lactase activity (45% higher, p<0.05). However, jejunal sucrase and maltase activities were not different (p>0.05) among different groups. The mice in LPS group exhibited lower level of DAO and ITP as well as higher TNF-╬▒ than did the Control groups (Table 3). Correspondingly, significantly improved DAO activity (61.48% higher, p<0.05) and ITP (17.75% higher, p<0.05) as well as reduced concentration of TNF-╬▒ (22.28% lower, p<0.05) were observed compared with those in LPS group. There were no differences on these indices between control and ASPS+LPS group (p>0.05).
Intestinal HSP70, EGF, and EGFR mRNA expression
The mice in LPS group displayed significantly lower EGF and EGFR gene mRNA expression as well as increased HSP70 gene expression in intestinal mucosa compared to those in control group (Table 4). In contrast to those in LPS group, the mice treated with ASPS had significantly raised mRNA expression of EGF (100% higher, p<0.05) and EGFR (200% higher, p<0.05) gene, and reduced the HSP70 (77.42% lower, p<0.05) gene expression. There were no differences on these indices between control and ASPS+LPS group (p>0.05).
DISCUSSION
ASPS has demonstrated its immune-regulatory and inhibitory inflammatory activity in animal models and clinical trials in many studies (Fang et al., 1985; Zhang et al., 1993; Steinmann et al., 2001; Han et al., 2003; Chen et al., 2011; Han et al., 2014). However, focusing on the gastrointestinal tract mucosa constantly being confronted with a variety of potentially injurious agents, few researches were conducted to determine the role of pre-treatment with ASPS on the intestinal mucosal barrier under inflammatory conditions.
In the current study, to evaluate whether ASPS pre-treatment could attenuate the damage to intestinal mucosal function in a challenged animal, we employed a well-documented model of mucosal impairment in mice built by administrating Escherichia coli LPS. This toxin molecule is present on the membrane of gram-negative bacteria and causes acute systemic inflammatory symptoms characterized by transient fever, anorexia, somnolence and inactivity (Spurlock, 1997), as well as intestinal morphologic alterations. Exposure to LPS further results in dysfunction of intestinal mucosa including negative alternations in digestive function, mucosal integrity and immune function (Kruzel et al., 2000), and has been frequently applied in many researches for studying nutrition and gastrointestinal physiology (Liu et al., 2008; Hou et al., 2010; Liu et al., 2012). In addition, the time point of sampling in the present experiment was chosen at 4 h post-injection with LPS as previous studies suggest that the acute intestinal inflammatory response occurs 3 to 6 h following LPS injection (Mercer et al., 1996; Liu et al., 2008; Liu et al., 2012).
Intestinal epithelial barrier consists of the intestinal morphology, which includes a tight junction (TJ) between epithelial cells, mucus layer overlying the mucous epithelium, various digestive enzymes that simultaneously affects biological and immunological barriers and plays vital role in protecting gut against a variety of injurious agents. Villus height, crypt depth, as well as VCR are the critical factors of the intestinal epithelial barrier and vital to the full digestion and absorption of nutrients (Scharek et al., 2005). Of them, enhanced villus height and VCR present absorption ability improvement through increased contact area, and decreased crypt depth reflects the elevated proliferation rate of IEC. Our present study demonstrated intestinal morphology damage at 4 h following LPS challenge. As expected, ASPS treatment resulted in the alleviation of intestinal morphology damage induced by LPS challenge which suggests potently that ASPS might improve intestinal nutrients absorption ability, and the reason may be partially do with the capacity of ASPS to sustain growth and differentiation of epithelial cells. However, the underlying mechanism needs further research.
The complete TJ between IEC is a vital structural foundation of the intestinal mucosal epithelial barrier (Vicente et al., 2001). In agreement with many researches demonstrating LPS-induced inflammation results in expression alterations of TJ proteins (Weiler et al., 2005; Liu et al., 2012), our study found that occludin-1 protein expression was reduced by LPS challenge and ASPS facilitated its protein expression to nearly restore it to control level. Therefore, our finding concerning improved intestinal occludin-1 protein expression suggests that ASPS can be a preventive nutrient to lessen the loss of the epithelial barrier suffering from inflammatory challenge.
Disaccharidases including lactase, sucrase, and maltase overlying the mucosal surface play a vital part in decomposing disaccharides and facilitating mucosa maturation and gut digestive function (Lorenzsonn et al., 1987). In the present study, ASPS increased intestinal lactase activity in challenged mice, which indicates that ASPS protected the intestinal digestive function from damage. Zheng et al. (1999) has reported that insulin can promote mucosa maturation, and increase mucosal disaccharidase activities in neonatal pigs. Interestingly, although not direct observation of insulin level, based on our previous research demonstrating that ASPS can induce the increase in growth hormone, insulin-like growth factor I and decrease glucose in LPS-challenged weaned piglets (Han et al., 2014), we can infer an increase in insulin concentration was induced by ASPS. Therefore, it is possible that ASPS supplementation attenuated the decrease in disaccharidase activity partially by stimulating insulin release.
DAO as intracellular enzyme with high activity exists in the intestinal villous cells and its activity declines when IEC is injured. Thus, the DAO activity of intestinal mucosa can reflect the changes in intestinal mucosa integrity, permeability and the barrier function (Zhang et al., 2002). In the current study, our findings concerning oral ASPS alleviated the loss of DAO activity further prove the intestinal mucosal function is boosted with the treatment of ASPS under the condition of challenge.
The modulation of mucosal inflammatory response by ASPS was assessed under inflammatory intestinal conditions resulting in compromised barrier function (Irvine and Marshall, 2000). Over-release of TNF-╬▒ to the mucosa induced by LPS is primarily responsible for the disruptive intestinal mucosal function (Pi├® et al., 2004; Field et al., 2010). Similar to TNF-╬▒, a high level of HSP70 is also indicative to inflammatory response in the intestine. Consistent with our previous study demonstrating the relief of inflammatory response by ASPS via modulating the release of pro-inflammatory cytokines including TNF-╬▒, interleukin (IL)-1╬▓, and IL-6 in the blood of piglets (Han et al., 2014), our current determination of inflammatory mediators showed that ASPS decreased the intestinal mucosal concentration of TNF-╬▒ and HSP70, which further support the above results concerning an improved intestinal mucosal barrier by ASPS.
It is apparent that the intestinal mucosal barrier is dependent on the presence of a monolayer of epithelial cells joined by TJ, which form a cellular barrier to many of the wide range of potential injuries. Although intestinal epithelium represents the cellular frontier of the mucosa, it has long been recognized that the apical surface of this population is covered by a viscoelastic coat including large amounts of trefoil peptides selectively secreted by goblet cells (Pi├® et al., 2004), which are in conjunction with mucin glycoproteins to form a continuous gel on the luminal surface to prevent intestinal barrier damage (Hoffmann, 2005). Previous studies revealed the ability of ITP in facilitating restitution of wounded mucosa and protecting the integrity of model intestinal epithelial monolayers against a variety of injurious agents (Dignass et al., 1994). In our current study, relative to lower number of goblet cells and ITP concentration in challenged mice, the mice in ASPS groups presented a higher number of goblet cells and concentration of ITP even when compared with control group, which may be due to the capacity of ASPS functioning under both challenged and normal conditions as reported in our previous study (Han et al., 2014). This observation also elucidates the distinct protective role of ASPS against injurious agents via promoting the formation of viscoelastic gel overlying the mucosa by the secreted products of the goblet cell population. However, the underlying mechanism needs further research.
EGF and its receptor EGFR are implicated by many studies (Barnard et al., 1995; Scharek et al., 2005) in the regulation of intestinal epithelium growth and differentiation, which are required for normal gut development and repair of intestinal mucosal injury (Abud et al., 2005). In the current study, these two indicators were examined to seek to further clarify the mechanism by which ASPS protect against mucosal barrier loss. EGF combination with its receptor EGFR conducts its stimulation signal intracellularly and can launch a series of cascade reaction through signaling pathways, where they act on transcription factors to modulate various physiological process (Park et al., 2005). As expected, the sharply down-regulated EGF and EGFR mRNA expression following LPS challenge indicate the presence of intestinal epithelial damage. However, for the mice given LPS, the findings of ASPS pre-treatment elevating mRNA expression of EGF and EGFR suggest that ASPS contributes to proliferation and differentiation of the IEC to a certain extent, which coincided with the positive alteration in intestinal morphology, TJ, and mucus layer. Especially, our finding that an increased spectrum of EGFR superior to those of EGF was observed, this may involve various growth factors participating in the mucosal developmental process where these growth factors compete to combine with EGFR and lead to increased need of EGFR. These results propose that improvement in intestinal epithelium growth and differentiation may be the mechanism by which intestinal mucosal barrier damage is ameliorated by ASPS.
IMPLICATIONS
The present study demonstrates that oral pre-treatment of ASPS could mitigate intestinal mucosal barrier loss under challenged conditions by modulating the mRNA expression of EGF and EGFR. These results suggest that improvement in intestinal epithelium growth and differentiation may be the mechanism by which damage to the intestinal mucosal barrier by challenging agents is ameliorated by ASPS.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







