



INTRODUCTION
Annual production of the Pacific oyster, Crassostrea gigas, in Korea, is 3.5 million ton, which occupies 90% of the total shellfish production (MIFAFF, 2012). Production of diverse lines through a breeding program and management of mother shell for the production of artificial seeds are essential to increase volume and quality of the oysters. As in other aquatic species, one of the most important traits in Pacific oyster aquaculture is fast growth rate, even after the introduction of triploid techniques (Guo and Allen, 1994) caused increase of production volume. In addition to the growth enhancement, traits responsible for disease resistance, e.g. to Haplosporidium nelson in eastern oysters, Crassostrea virginica (Haskin and Ford, 1979; Allen Jr., 1998), resistance to Bonamia in European fat oysters, Ostrea edulis (Naciri-Graven et al., 1998) and resistance to Marteilia sydneyi in Sydneyrock oyster, Saccostrea glomerata (Nell and Perkins, 2006) have been successfully introduced. Recently, a color factor, i.e. shell and mantle colors, has been regarded as a new high potential trait for a better commercial value (Ward et al., 2000; Brake et al., 2004).
It is known that color of seafood plays a key role for consumer choice because it influences taste thresholds, sweetness perception, food preference, pleasantness, and acceptability (Clydesdale, 1993). For example, consumers are willing to pay more for the rich red salmon fillets (Steine et al., 2005; Alfnes et al., 2006). The shell and mantle color of the Pacific oyster also influence consumer preferences of oysters. For example, some “golden” oysters, displaying an orange/bronze or “golden” mantle and shell, have been developed and maintained for the color despite slow growth of the oysters (Nell, 2001). The mantle color of the Pacific oyster is, therefore, one candidate trait for the Molluscan Broodstock Program (MBP) of the United States (Brake et al., 2004). In Korea, the Pacific oyster with black mantle are favored by consumer and traded at about 20% higher price. However, no strain or line with black mantle has been yet established by systematic breeding plans.
Selection has been successfully implemented for the color manipulation of edible and ornamental aquaculture species (Gomelsky, 2011). Some color traits of aquatic animals are inherited either by qualitatively (non-additive) or quantitatively (additive). Examples of non-additive inherited color traits are pigmentation in rainbow trout, red and white color patterns in koi carp, and color in Tilapia sp. (Thorgaard et al., 1995; Gomelsky et al., 1996; Lutz, 2001). In contrast, the color traits of coho salmon, Atlantic salmon, and rainbow trout are known to be inherited in additive fashion (Gjerde and Gjedrem, 1984; Gjerde and Schaeffer, 1989; Withler and Beacham, 1994).
There are few studies on genetics of shell or mantle pigmentation of Pacific oysters. Brake et al. (2004) reported positive correlation between pigmentation of shell and mantle edge. Evans et al. (2009) reported that shell pigmentation was an inheritable trait in a continuous distribution fashion in most of the twenty-six full-sib families that were derived from parents collected from a naturalized population.
In this study, F1 and F2 families of the Pacific oyster were produced by crossing F0 parents with black and white shell pigmentations, in order to characterize distribution patterns and relationship of colors between shell and mantle edge, and to estimate heritability of the two traits, which can provide valuable information for the efficient breeding of shell and mantle edge colors in Korean oyster populations, where various types of the two phenotypes exist.
MATERIAL AND METHODS
Data collection
Parents were chosen from six aquaculture farms in Tongyeong, Gyeongsangnam-Do, Korea. The parents were produced in 2010 by collecting naturally produced larvae and cultured in aquaculture farms using a hanging method of seeded culture. Six F1 families were generated by single-pair mating with the greatest scores for each color, i.e. black (male)×black (female) or white (male)×white (female) cross. The larvae were settled onto cultch and cultured as described above. Among the F1 individuals, siblings with the greatest scores for each color were chosen as F1 parents at the age of 12 months, such that two and four F2 families were generated by black×black and white×white F1 cross, respectively. For the mating, naturally matured gametes were obtained by strip-spawning and consequently fertilized. Each family was kept in separate tank and seeded on different rope for hanging culture. After 5 month culture, 30 to 50 individuals from each F1 or F2 family were measured for shell color and mantle edge pigmentation; 45, 32, and 39 for black F1, 54, 51, and 44 for white F1, 34 and 51 for black F2, and 51, 50, 50, and 50 for white F2 families, respectively. A control sample of F0 240 individuals that were collected from six aquaculture farms in Tongyeong area in 2011 was also used to measure shell color and mantle edge pigmentation.
Measurement of shell and mantle edge pigmentation
Shell color and mantle edge pigmentation were classified into six and ten levels, respectively, depending on magnitude of darkness. The category of shell color was similar to Imai and Sakai (1961) and Brake et al. (2004), in which shell pigmentation was classified into five and four levels, respectively. However, six levels of shell color were applied in this study for better discrimination of color variation. The categories of the shell pigmentation were as follow:
Left and right shells are all white, no pigmentation present - score 0,
One shell is white and the other shell is pigmented over 50% - score 1,
Left and right shells are pigmented less 50% - score 2,
One shell is white and the other shell is pigmented over 80% - score 3,
One shell is pigmented less 50% and the other shell over 80% - score 4,
Both the left and right shells are pigmented over 80% - score 5.
The mantle edge pigmentation were scored from 1 to 10 by eyes according to the neutral color scheme that represented snow white, pearl white, silver gray, metal gray, aluminum gray, slate gray, charcoal gray, carbon black, and real black, respectively.
Data analysis and estimation of heritabilities
The data for shell color and mantle edge pigmentation were analyzed to test any phenotypic difference between the two colors (black and white) in F1 or F2 generation. The following linear model was used:
Where Yijk represent measurement of pigmentation (shell or mantle edge) of kth individuals in jth family within ith color (1: black, 2: white), Ci represent the fixed effect of color i, Famj(Ci) represent the random effect of jth family nested within ith color, eijk represent the residual of kth individual of jth family within ith color. The model was analyzed using SAS GLM procedure (SAS Institute, 2000, Cary, NC, USA), and variance components of the residual (σ2e) and family (σ2F) effects were estimated using SAS PROC VARCOMP option. Estimates of the variance components were obtained for each F1 and F2 generation. The additive genetic variance (σ2g) could be estimated under a full-sib family structure (Falconer and Mackay, 1996) as σ2g = 2σ2F, and a narrow sense heritability (h2) was obtained as h2 = σ2g/(σ2g+σ2e). Spearman correlation coefficients between phenotypes of shell color and mantle edge pigmentation were also calculated using SAS PROC CORR option.
RESULTS
Distribution of F0, F1, and F2 oysters for shell and mantle edge pigmentation
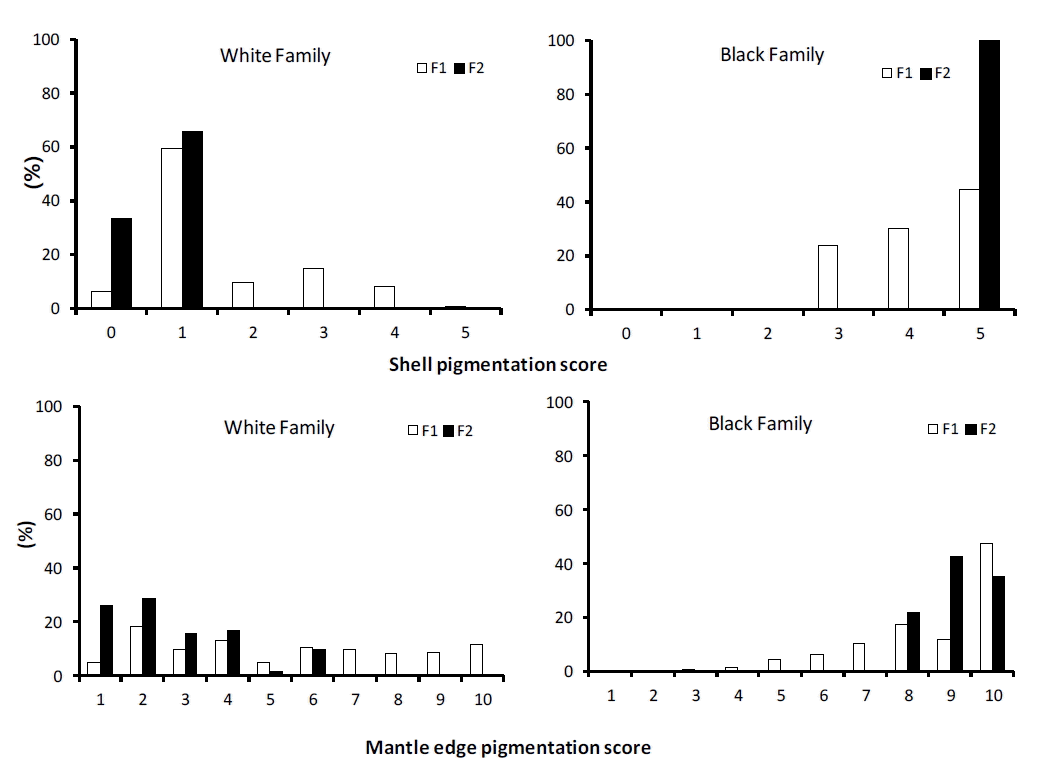
Among the 240 F0 individuals, proportion of the oysters with black shell (level 4 and 5) and black mantle edge (levels 8 to 10) was 29.2%, 10.8% with white shell (level 0 and 1) and white mantle edge (levels 1 to 3), 8.1% with white shell and black mantle edge, and 1.3% with black shell and white mantle edge, respectively. The rest (50.6%) of the F0 oysters had shell colors with level 2 or 3 and mantle edge colors with level 4 to 7. The overall mean and standard deviation of shell color and mantle edge pigmentation in F0s were 2.59±1.55 and 6.33±2.30, respectively, ranging 2.27 to 3.07 for shell color and 5.30 to 7.70 for mantle edge pigmentation, respectively. The Pearson correlation coefficient between shell color and mantle edge pigmentation in the F0 individuals was 0.25 (p<0.0001).
Means and standard deviations of shell color and mantle edge pigmentation in F1s and F2s are described in Table 1. In the black cross families, the overall means (standard deviation) were 4.19 (0.83) and 5.00 (0.00) for shell color, and 8.60 (1.70) and 9.20 (0.77) for mantle edge pigmentation in the F1 and F2 generations, respectively. Especially, all F2 individuals in the black cross families had black shell color with the maximum level of five, i.e. with zero standard deviation. In contrast, in the white cross families, the average values of shell color were 1.60 (1.11) and 0.67 (0.48) in the F1s and F2s, and 5.40 (2.88) and 2.72 (1.57) for mantle edge pigmentation in the F1s and F2s, respectively. Proportions of oysters with black shell (level 4 and 5) were 75% and 100% in the F1 and F2 black cross, and 67% and 100% with white shell (level 0 and 1) in the F1 and F2 white crosses families, respectively (Figure 1). Similarly, proportions of offspring with black mantle (level 9 and 10) were 59% and 79% in the F1 and F2 black cross, while 23% and 55% of oysters had white mantle edge (level 1 and 2) in the white F1 and F2 cross families, respectively (Figure 1). These results show that the frequencies of individuals with black (white) shell and black (white) mantle edge in black (white) cross F2 families were greater (smaller) than in the F1 families with the corresponding colors (Figure 1). Also, standard deviations of the two color traits in the F2s were smaller than in the F1s, indicating that F2 individuals had more homogeneous colors of shell and mantle edge than the F1s in the black or the white crosses (Table 2). Also, the phenotypic relationship between shell color and mantle edge pigmentation was greater in the F2s than in the F1s, i.e. the Spearman correlations between the two traits were 0.92 and 0.74 in the F2 and F1 generations, respectively (p<0.0001).
Analysis of variance and estimation of heritability
The effects of color and families nested within color on shell and mantle edge pigmentation were significantly different (p<0.05) in both F1 and F2 generations (Table 2). The effects of color or family were much more significant in F2 than in F1, expect for the family effect of mantle edge color. This may be partly due to smaller estimates of the error (residual) variances in F2s.
Estimates of variance components showed that the residual, family (genetic) and phenotypic variances in F2 generation were smaller for both shell and mantle edge colors than in F1 (Table 3). Estimates of heritability were 0.41 and 0.77 for shell color and 0.27 and 0.08 for mantle edge pigmentation in F1 and F2 generations, respectively.
DISCUSSION
The Pacific oyster with black mantle edge is favored by Korean consumers. However, specific lines for the trait were not developed, because the genetics of this pigmentation has not been studied, let alone genetic relationship of the trait with shell pigmentation. Although, the relationship between shell and mantle pigmentation (Brake et al., 2004), and heritability of shell pigmentation (Evans et al., 2009) in Pacific oyster were reported, the results need to be verified for the oysters in Korea. In this study, we produced two generations of the Pacific oyster from a random oyster sample population by crossing individuals with black or white color for both shell and mantle edge, and analyzed the phenotypes to characterize relationship between shell and mantle edge pigmentation and to estimate heritabilities of the two color traits.
Analysis of a natural population (F0), from which F1 parents were produced, showed that pigmentation of shell and mantle edge had various patterns. Among the F0s, 40% of individuals had consistent colors, i.e. with black or white for both shell and mantle edge, while half (50.6%) of the F0 individuals had pigmentation of medium level for the two color traits. However, the individuals with black shell and white mantle edge represented only 1.3% (3 out of 240). These results suggest that oysters with black (white) shell have more tendency to have black (white) mantle, as reported by Brake et al. (2004). In fact, the oysters with black shell and black mantle edge or white shell and white mantle edge were 29.2% and 10.8% of the F0 population, respectively (data not shown).
Intrigued by the results, individuals with black or white color for both shell and mantle edge were selected and crossed for two (F1 and F2) generations. Analyses of shell and mantle pigmentation of the F1 and F2 families showed that there were increasing proportions of individuals with the same color for both shell and mantel edge with generation, i.e. more F2s had black (white) color for both shell and mantle edge than the F1s in black (white) cross families (Table 1). These results suggest that selection based on shell and mantle edge color is effective in the Pacific oyster in Korea, and that the two colors are heritable traits, which were supported with the heritability estimates (Table 3). Spearman correlation coefficients between shell and mantle edge color were 0.25, 0.74, and 0.92 in F0, F1, and F2 generations, respectively, indicating that the relationship between the two colors were stronger in successive generations. Brake el al. (2004) also found the positive correlation (r = 0.578) between the two color traits in the Pacific oyster, i.e. the correlation was statistically significant in two of the three cohorts, in which selection programs for growth were implemented. This result strongly suggest that selection considering both for shell and mantle edge color is very effective, and thus, with generations, an individual with black (white) shell color is more likely to have black (white) mantle edge pigmentation.
Although shell pigmentation had various color patterns, indicating polygenic nature of the trait (Evans et al., 2009), there are evidences that a few major genes are involved in the determination of shell pigmentation. Imai and Sakai (1961) reported that hybrid offspring from the crosses between light and dark strains had intermediate shell pigmentation. Among the 26 full-sib oyster families analyzed by Evans et al. (2009), the shell color had non-normal distributions in eight families and offspring segregation ratio of 3:1 for lighter and darker shell groups was observed in another two families. Hedgecock et al. (2006) reported that a single additively-inherited QTL could explain 32% of the phenotypic variation in total left-shell pigmentation in an experimental population derived from two partially inbred lines. Thus, QTL or other molecular studies are needed to characterize genetic architecture of the shell and mantle edge pigmentation in the Pacific oyster in Korea.
Variance component estimates showed that the estimate values of residual (environment), family (genetic) and phenotype were smaller in F2 than in F1 generation for both shell and mantle edge pigmentation (Table 3). This may partly be due to selection process, i.e. as selection for the color traits was implemented, the individuals within each color cross family would become more homogenous in the next generation, such that genetic variance would become smaller. However, the heritability estimates of mantle edge color decreased, while those of shell color increased as generation went on (Table 3). Further study with an increasing sample size is needed to obtain more reliable estimates of the variance components and thus heritability.
Our results indicate that some proportions of phenotypic variations for the shell and mantle edge colors were explained by genetic variations, enabling effective breeding implementation for the color traits. Because color patterns of aquatic species are often influenced by environmental factors, further studies are needed to investigate how environmental factors influence pattern distributions of the shell and mantle edge pigmentations, as well as to confirm the results of genetic characterization of the color traits in the Pacific oyster in this study.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







