



INTRODUCTION
Similar to interferon alpha (IFNA) gene family, interferon tau (IFNT), which encodes a trophoblast factor essential for the process of maternal recognition of pregnancy in ruminant ungulates, belongs to a gene family. Exhaustive experiments have been conducted to characterize the effects of IFNTs, but IFNT paralogs actually expressed in utero and their effects on uterine epithelial cells have not been carefully evaluated.
In mammalian females, the maintenance of corpus luteum function beyond the length of the normal estrous cycle and the continued secretion of steroid hormones are required to establish and maintain pregnancy. Pregnancy recognition signaling in domestic ruminants (Short, 1969) is a local phenomenon involving a conceptus factor that acts on the uterine endometrium to alter gene expression (Spencer and Bazer, 2004). During the peri-implantation period, the embryonic trophectoderm of the ruminant conceptus secretes the cytokine IFNT (Imakawa et al., 1987; Roberts et al., 1992), which acts on the uterine endometrium (Godkin et al., 1984) and attenuates endometrial production of the luteolysin prostaglandin F2α, in turn maintaining corpus luteum function (Spencer et al., 1996; Salamonsen et al., 1998). Secretion of ovine IFNT starts on d 8 to 9 of pregnancy (d 0 = day of estrus), peaks on d 16 to 17, and then declines rapidly (Godkin et al., 1982; Ashworth et al., 1989; Farin et al., 1989). Thus, IFNT expression is limited to the peri-attachment period, during which the blastocyst begins to elongate, while up to approximately 20% of trophoblast cells form binucleate cells (Xie et al., 1996; Klisch et al., 2006) and attach to the uterine epithelium.
A number of IFN-stimulated genes (ISGs), including the MX genes, are expressed in the uterus (Spencer et al., 2007; 2008). These ISGs are hypothesized to regulate endometrial receptivity to implantation as well as survival, growth, and development of the conceptus. In fact, ISG and MX are up-regulated in the ovine uterus in response to IFNT (Charleston and Stewart, 1993; Ott et al., 1998; Johnson et al., 1999). During the course of our studies, a next-generation sequencer was used to survey transcripts found in bovine conceptuses during the peri-implantation period (unpublished observation). To our surprise, only 2 IFNT transcripts, IFNT1 and IFNTc1, were found in the bovine uterus. However, uterine response to each of these IFNTs has not been executed.
The aim of this study was to characterize the response patterns of ISG12, ISG15, MX1, and MX2 after exposure to bovine IFNs (bIFNs)-IFNT1, IFNTc1, and IFNA-in uterine epithelial and Mardin-Darby bovine kidney (MDBK) cells. We determined that the expression of ISG12, ISG15, MX1, and MX2 transcripts in the bovine cells was influenced by recombinant bIFNs (rbIFNs) in a time- and concentration-dependent manner.
MATERIAL AND METHODS
Cell culture
Bovine uterine epithelial cells were kindly provided by Dr. K. Okuda, Okayama University. Briefly, cells were cultured in a growth medium composed of Dulbecco’s modified Eagle medium (DMEM)/nutrient mixture Ham’s F12 and RPMI medium 1640 (Sigma-Aldrich, St. Lois, MO, USA) containing 5% fetal bovine serum (FBS; JRH Biosciences, Lenexa, KS, USA) and antibiotics (Sigma-Aldrich) at 37°C in a 5% CO2 incubator. MDBK and human 293 cells (HTB36, ATCC) were grown in DMEM supplemented with 10% FBS (JRH Biosciences) and antibiotics at 37°C in 5% CO2.
Construction of bIFNs subtype vectors for recombinant protein production
Full-length complementary DNA (cDNA) regions of bIFNT1, bIFNTc1, and bIFNA (GenBank accession Nos. bIFNT1: M60903; bIFNTc1: AF238613; and bIFNA: NM_001017411) were amplified via polymerase chain reaction (PCR) using specific primers (Table 1). Each of these products was inserted into mammalian expression vector pcDNA3.1/v5-His-TOPO (Invitrogen, Carlsbad, CA, USA). Each vector was completely sequenced to rule out the possibility of PCR errors.
Production and purification of recombinant bIFNs
The plasmid was transfected into 293 cells using HilyMax transfection reagent (Dojin Chemicals, Kumamoto, Japan) following the manufacturer’s instructions. After 48 h, the cells were subjected to selection for stably transfected cells using 400 mg/ml G418. Selection was continued until monolayer colonies formed. The transfectants were transferred and maintained in DMEM supplemented with 10% FBS and 400 mg/ml G418. After the transfectants were grown to 90% confluence, the medium was discarded, and fresh DMEM medium containing 1.5% FBS, 400 mg/ml G418, and 15 mM sodium butyrate was added. One day later, the culture medium was collected. Target proteins from the culture medium were purified using His-tagged protein purification reagent (Medical and Biological Laboratories, Nagoya, Japan) according to the manufacturer’s protocol.
Western blot analysis
Western blot analysis was performed using the culture medium and purified recombinant protein. Protein concentrations were determined using a protein assay dye reagent concentrate (Bio-Rad Laboratories, Hercules, CA, USA). Cell lysates (10 μg) were loaded into each lane and separated with 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and then transferred onto a polyvinylidene fluoride membrane (Millipore, Bedford, MA, USA) and treated with rabbit anti-bIFNT antibody (Protein Purify, Isesaki, Japan) and rabbit anti-His-probe antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) diluted to 1:1000 with phosphate-buffered saline. The proteins were detected using secondary antibody conjugated with horseradish peroxidase and an enhanced chemiluminescence western blotting detection system (Amersham Pharmacia Biotech, Buckinghamshire, UK). Normal rabbit immunoglobulin G (IgG; Sigma-Aldrich) was used as a negative control.
rbIFN treatment on uterine epithelial and MDBK cells
Bovine uterine epithelial cells and MDBK cells were treated with the rbIFNs (1, 5, and 10 μg/ml) in 3 replicates, each to determine their response to different concentrations of each protein. Cells were isolated at 24, 48, and 72 h after treatment for the evaluation of response patterns of ISG12, ISG15, MX1, and MX2 messenger RNAs (mRNAs).
RNA extraction and reverse transcription (RT)-PCR
Total RNAs were extracted from each sample using Isogen reagent (Nippon Gene, Tokyo, Japan) according to the manufacturer’s protocol (Nagaoka et al., 2003). After extraction, the RNA was subjected to reverse transcription (RT) into cDNA using oligo (dT) 12 to 18 primers and SuperScript II (Gibco BRL Life Technologies, Rockville, MD, USA) according to the protocol suggested by the manufacturer. The cDNA molecules were used as templates for PCR analysis. Levels of ISG12, ISG15, MX1, and MX2 were examined via PCR using specific primers (Table 2). The PCR mixture consisted of 1 μl RT product, 1 μl 10×PCR buffer, 0.4 μl each forward and reverse primers (10 pM), 0.2 μl deoxyribonucleotide triphosphate mixture (10 mM), 0.3 μl MgCl2 (50 mM), 6.6 μl double-distilled H2O, and 0.1 μl Taq DNA polymerase (5 U/μl; Invitrogen). The PCR was performed under the following conditions: 94°C for 5 min, followed by 30 cycles of 94°C for 1 min, 57°C for 1 min, and 72°C for 1 min. The PCR product was analyzed via electrophoresis on 1% agarose gel stained with ethidium bromide. β-actin (ACTB) mRNA was used as an internal control.
Statistical analysis
RT-PCR data were analyzed using a Chemi-imager with AlphaEase software (Alpha Innotech Corp., San Leandro, CA, USA). Cells were the experimental unit and the fold changes of dependent variables were tested against concentration, time, and concentration×time. When significant concentration×time interaction was detected, variables were analyzed within the time. Data are presented as least squares means±standard error of the mean of relative fold change from 0 h levels using the critical threshold method.
RESULTS
Structural analysis of bIFNT1, bIFNTc1, and bIFNA and production of recombinant proteins
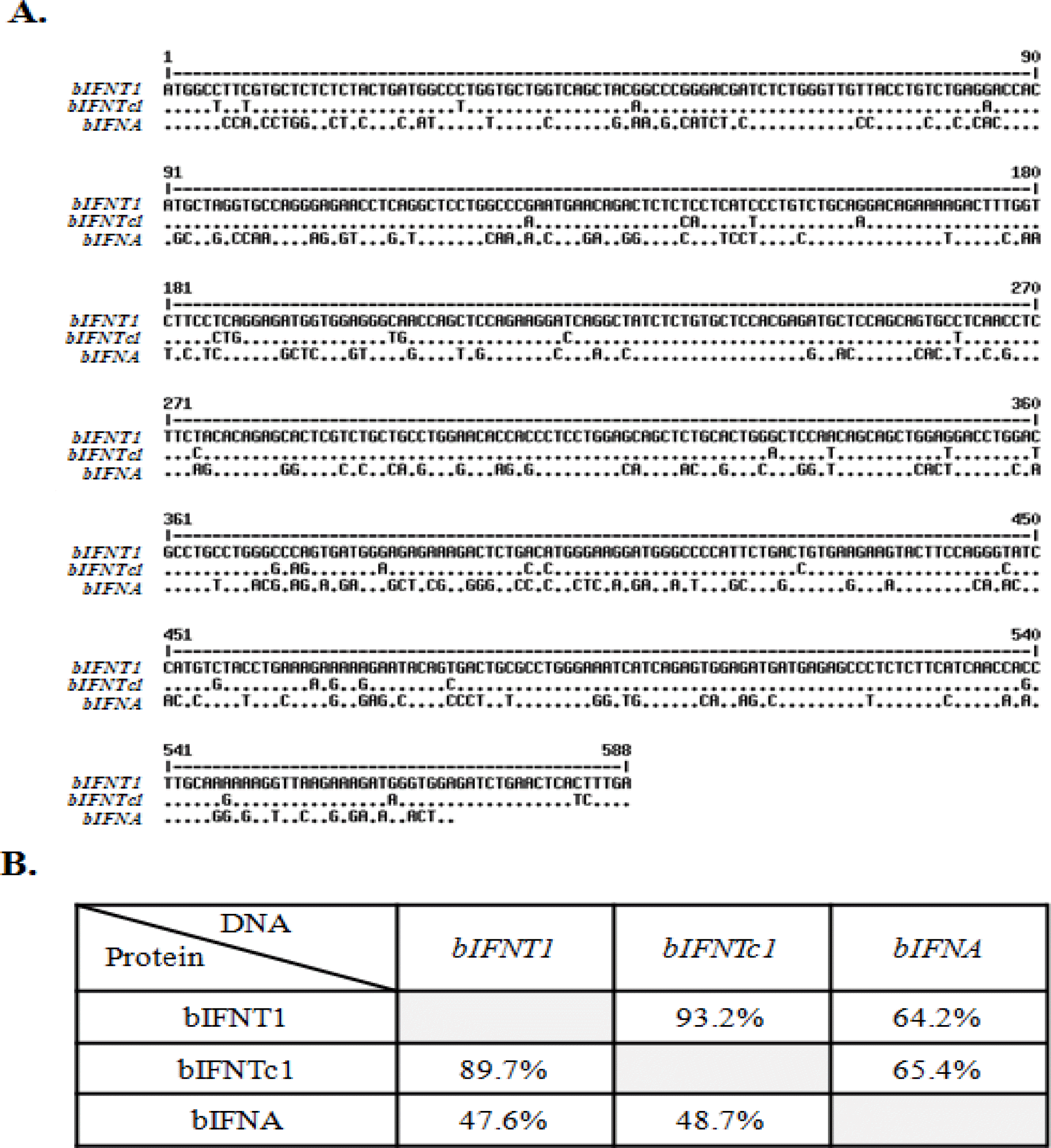
Comparison of the nucleotide sequences of the bIFN genes bIFNT1, bIFNTc1, and bIFNA is shown in Figure 1A. The homology search revealed that the nucleotide sequence of bIFNT1 had high homology with that of bIFNTc1 (93%). However, the sequence showed low homology with bIFNA cDNA (64%; Figure 1B).
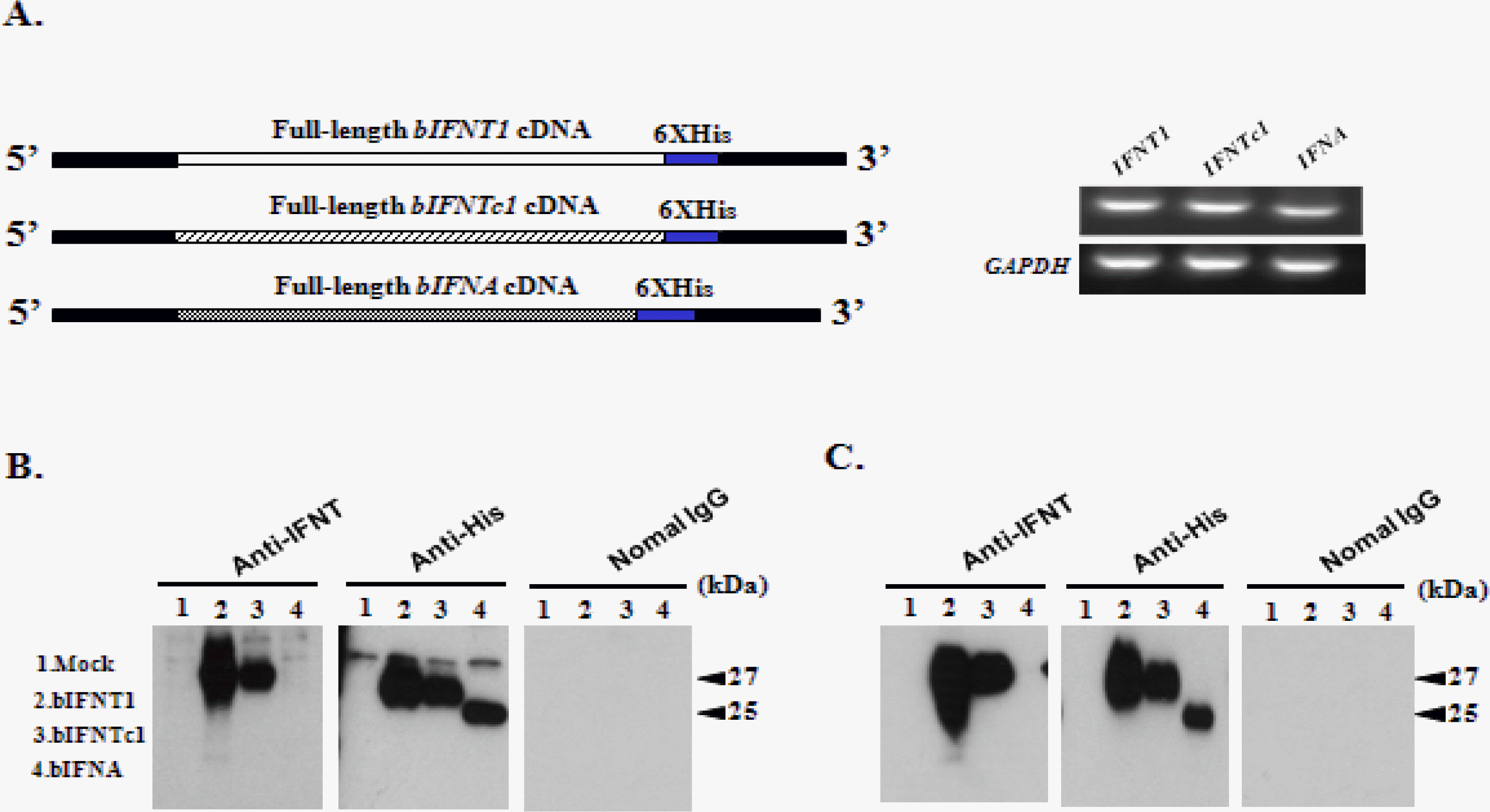
Three expression vectors (pcDNA3.1-bIFNT1, pcDNA3.1-bIFNTc1, and pcDNA3.1-bIFNA) were transfected into 293 cells (Figure 2A), from which culture media were collected and purified via a His-tagged protein purification method. The supernatant was subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Specific antibodies for IFNT and His were used to detect recombinant proteins produced by the 3 expression vectors via western blot (Figure 2B, C). rbIFNT1 and rbIFNTc1 showed single bands with molecular weights of 27 kDa. However, the band was a slightly lower at 25 kDa for bIFNA owing to shortening by 6 amino acids. No band was detected in the fresh medium or with normal IgG (Figure 2B, C). The 27 kDa band represented the approximate molecular weight predicted from the rbIFNT1 and rbIFNTc1 cDNA sequences, including the carbohydrate chains.
Up-regulation of ISG12 and ISG15 mRNAs by rbIFNs in uterine epithelial cells
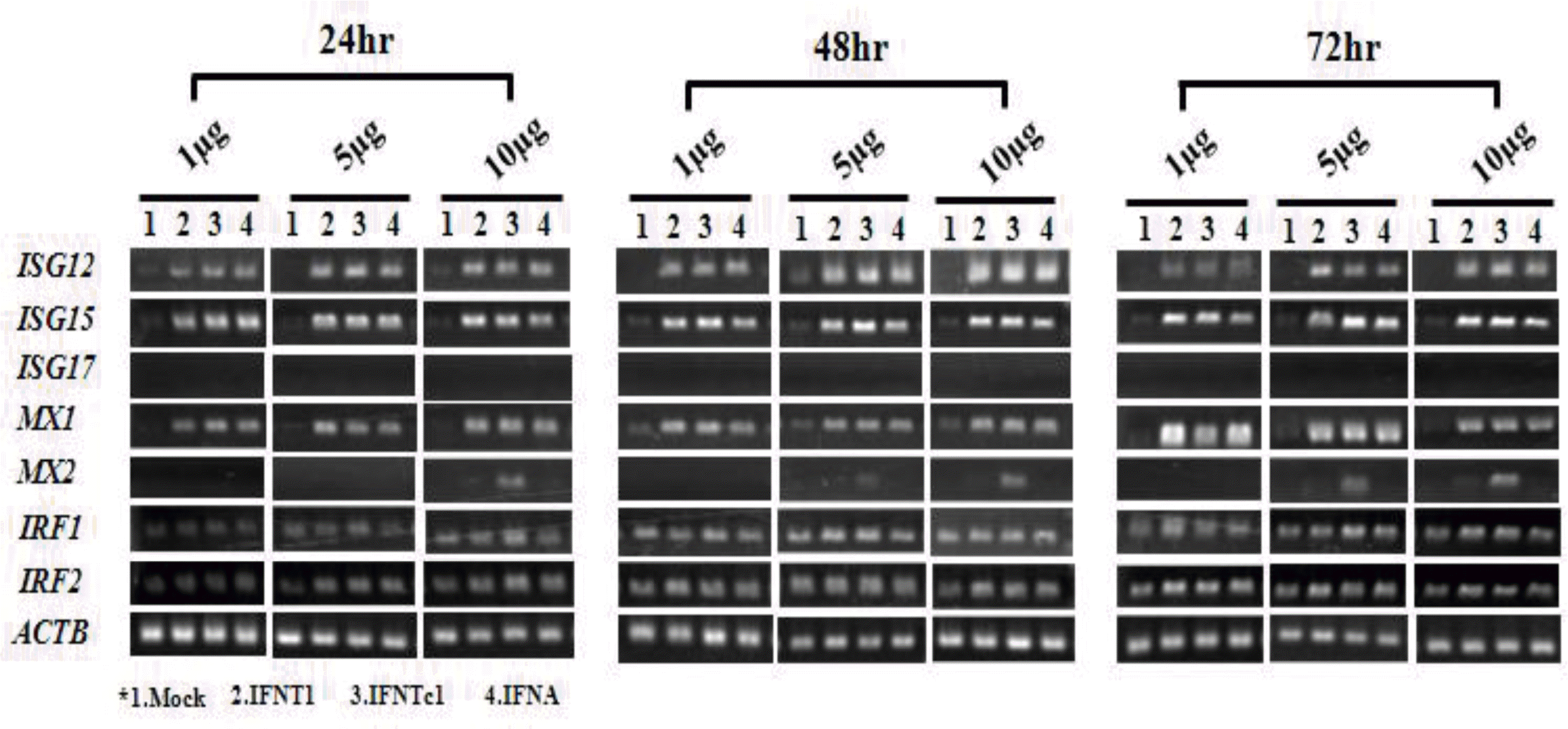
To investigate responsiveness to rbIFNs in uterine epithelial cells, ISG, MX, and interferon regulatory factor gene (IRF) transcripts were examined after rbIFN treatment. A concentration- and time-dependent response was recorded with 1, 5, and 10 μg/ml at 24, 48, and 72 h. Recombinant proteins used in this study affected ISG12 and ISG15 transcripts but not that of ISG17 (Figure 3). Likewise, these treatments had some effects on MX1 and MX2 transcripts but not on IRF transcripts (Figure 3). Therefore, ISG17 and IRF were excluded from subsequent quantitative studies. All rbIFNs significantly up-regulated ISG12 and ISG15 mRNAs after 24, 48, and 72 h of incubation. ISG12 mRNA was remarkably increased at 48 h. After that, it decreased to the same level as that measured at 24 h (Figure 4A). Stimulation by rbIFNs also significantly increased the expression of ISG15 mRNA. In particular, the ISG15 mRNA level was increased at 24 h after treatment and was maintained through 72 h (Figure 4B), and this effect was not dose dependent. In the rbIFNA treatment, the expression levels of ISG15 mRNA were significantly decreased at 48 h.
Changes in MX1 and MX2 mRNAs by rbIFN stimulation in uterine epithelial cells
rbIFNs significantly up-regulated the level of MX1 mRNA at 24, 48, and 72 h after treatment (Figure 4C). The up-regulation of MX1 was mild in the first 24 h post-induction with rbIFNT1, rbIFNTc1, and rbIFNA (1 and 10 μg/ml), then slightly decreased at 48 h, and was increased at 72 h. The MX2 expression levels gradually increased and peaked at 72 h after treatment with 10 μg/ml rbIFNTc1 and showed a high increase only at 72 h (Figure 4D). MX2 mRNA was not affected by rbIFNT1 or rbIFNA in the uterine epithelial cells. Thus, the patterns of MX1 and MX2 responses to rbIFNs were different.
Up-regulation of ISG12 and ISG15 mRNAs by rbIFNs in bovine MDBK cells
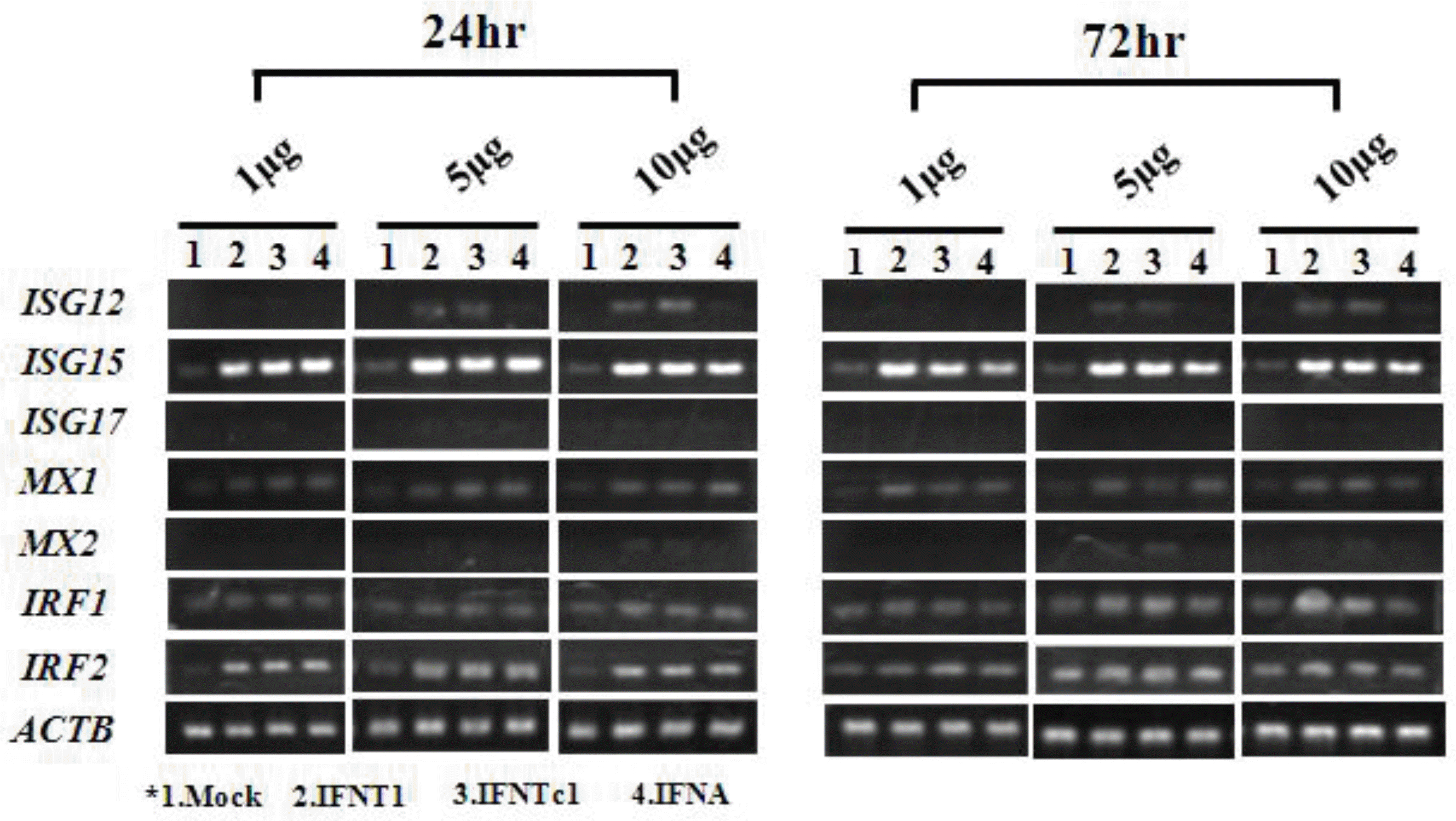
rbIFNs significantly up-regulated ISG12 and ISG15 mRNAs at 24 and 72 h after treatment in bovine MDBK cells (Figure 5), and the expression was dramatically increased in the rbIFNT1 and rbIFNc1 treatment groups. ISG12 mRNA expression was unchanged by rbIFNA treatment (Figure 6A), but ISG15 mRNA expression was significantly increased at 24 and 72 h after rbIFNA, rbIFNT1, or rbIFNTc1 treatment (Figure 6B). Thus, the mRNA expression pattern for ISG15 was similar in uterine epithelial cells and MDBK cells. However, ISG12 mRNA demonstrated a slightly different expression pattern after rbIFNA treatment.
Changes in MX1 and MX2 mRNAs by rbIFN stimulation in MDBK cells
MX1 mRNA expression increased at 24 h after rbIFNT1, rbIFNTc1, and rbIFNA treatment. However, it decreased slightly at 72 h after rbIFNA treatment. By contrast, MX1 mRNA expression increased slightly at 72 h after rbIFNTc1 exposure (Figure 6C). MX2 expression gradually increased after rbIFNT1 or rbIFNTc1 treatment, but it was unchanged by rbIFNA (Figure 6D). These patterns did not differ between uterine epithelial cells and MDBK cells.
DISCUSSION
IFNT from a blastocyst (d 15 to 16 after breeding in cattle) clearly alters endometrial gene expression, allowing the establishment of pregnancy in ruminant ungulates (Spencer and Bazer, 2004). To our knowledge, this is the first study to examine the effect of IFNT paralogs, of which transcripts have been found in the bovine uterus on endometrial epithelial cell responses. The examination of recombinant IFNs in this study revealed that IFNs have common functions as well as functions unique to each paralog. This observation further suggested that endometrial responses to IFNT must be re-evaluated using each IFNT isoform as well as with a combination of IFNT isoforms. Likewise, in vivo IFNT infusion study requires reevaluation based on the dose and kinds of IFNT isoforms, which may affect the outcome.
Until recently, IFNT was thought to act exclusively in a paracrine manner in the endometrium, causing endometrial responses in pregnant sheep and cattle (Vallet et al., 1991; Charleston and Stewart, 1993; Ott et al., 1998; Spencer et al., 1998; Johnson et al., 2000). ISGs are induced systemically within days of blastocyst signaling in ewes (Yankey et al., 2001). In addition, Oliveira and coworkers (2008) showed that ISG expression occurs in extrauterine tissues during early pregnancy, possibly owing to endocrine IFNT release from the uterine vein. These studies do not completely prove that IFNT could escape from the uterus and initiate a peripheral antiviral response. Regardless of proof of direct or indirect effects of IFNT in the circulation, the degree of response observed in ISGs and MXs in this study suggested that ISGs escaped from the uterus into the circulation.
Charleston and Stewart (1993) first showed that MX was strongly up-regulated in ovine endometrium during early pregnancy. Later, MX was shown to be expressed in the ovine uterus during the estrous cycle (Ott et al., 1998). The roles of ISG17 and MX in the ovine endometrium are unknown, but their temporal and spatial patterns of expression provide insight into their physiological relevance. One interpretation of these results from primates, rodents, ruminants, and pigs is that ISG17 and MX are part of a universal uterine response to fetal/placental development. In support of a general role for MX in early pregnancy, its expression has been shown in the uterine endometria of sheep (Ott et al., 1998), cattle, pigs, and horses (Hicks et al., 2003), and rodents (Chang et al., 1990). The regulation and immediate functional implications of these various expression patterns may be independent but are almost certainly coordinated to favor conceptus development and survival.
In summary, the results of this study indicate that rapid communication occurs between the conceptus and the uterine endometrium at approximately the time of maternal recognition of pregnancy in cattle. These changes result in gene expression that can be measured in uterine epithelial cells and possibly in the circulation. Uterine epithelial ISG12, ISG15, MX1, MX2 were activated, whereas IRF1 and IRF2 were not. The required adaptor molecules and transcription factors were mostly up-regulated in a time- and concentration-dependent manner. These results led us to conclude that bovine uterine epithelial cells respond to rbIFNs differently, which warrants further investigation.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







