- Invited Review - Calcium Digestibility and Metabolism in Pigs*
Article information
Abstract
Calcium (Ca) and phosphorus (P) are minerals that have important physiological functions in the body. For formulation of diets for pigs, it is necessary to consider an appropriate Ca:P ratio for an adequate absorption and utilization of both minerals. Although both minerals are important, much more research has been conducted on P digestibility than on Ca digestibility. Therefore, this review focuses on aspects that are important for the digestibility of Ca. Only values for apparent total tract digestibility (ATTD) of Ca have been reported in pigs, whereas values for both ATTD and standardized total tract digestibility (STTD) of P in feed ingredients have been reported. To be able to determine STTD values for Ca it is necessary to determine basal endogenous losses of Ca. Although most Ca is absorbed in the small intestine, there are indications that Ca may also be absorbed in the colon under some circumstances, but more research to verify the extent of Ca absorption in different parts of the intestinal tract is needed. Most P in plant ingredients is usually bound to phytate. Therefore, plant ingredients have low digestibility of P due to a lack of phytase secretion by pigs. During the last 2 decades, inclusion of microbial phytase in swine diets has improved P digestibility. However, it has been reported that a high inclusion of Ca reduces the efficacy of microbial phytase. It is possible that formation of insoluble calcium-phytate complexes, or Ca-P complexes, not only may affect the efficacy of phytase, but also the digestibility of P and Ca. Therefore, Ca, P, phytate, and phytase interactions are aspects that need to be considered in Ca digestibility studies.
INTRODUCTION
Although minerals are required in small quantities in diets for pigs, inorganic elements are essential for normal growth and reproduction in pigs. The concentration of minerals in the animal body is 2 to 5% depending on the species, and the skeleton system contains the largest amounts of minerals (Gillespie, 1987). Calcium (Ca) is the most abundant mineral in the body followed by phosphorus (P; Kellems and Church, 1998). Between 96 and 99% of the total Ca in the body and between 60 and 80% of total P in the body is stored in bone tissue (Crenshaw, 2001). Bone ash contains 36 to 39% Ca and 17 to 19% P (Crenshaw, 2001), and minerals are also present in soft tissue such as blood, body fluids, and some secretions, and they are involved in many biochemical reactions in the body (Gillespie, 1987; Kellems and Church, 1998). Calcium and P are also important for physiological functions such as muscle contraction, transmission of nerve impulses, enzyme activation, metabolic reactions, protein synthesis, maintenance of osmotic and acid-base balances, components in membranes, and other functions (Crenshaw, 2001; Ewing and Charlton, 2007).
Calcium and P are considered macrominerals because they are required at levels greater than 100 ppm in the diets (Ewing and Charlton, 2007). However, inclusion of each mineral must be controlled because an interaction exists between Ca and P, which may influence absorption of both minerals (Crenshaw, 2001; Ewing and Charlton, 2007). Excess or deficiency in one of the minerals causes impaired utilization of the other. The concentration of Ca in cereal grains, oil seed meals, and many other plant ingredients is very low compared with concentrations in animal proteins such as fish meal and meat and bone meal, or inorganic minerals such as limestone, calcium carbonate, and calcium phosphates. Plant ingredients have low bio-availability of P because some of the P in these ingredients is bound to phytate. Inclusion of microbial phytase in swine diets results in release of some of the P that is bound to phytate and improves P digestibility and reduces the excretion of P from the pigs (Cromwell et al., 1995; Almeida and Stein, 2010; Kerr et al., 2010). However, high levels of Ca in swine diets reduce the effect of phytase and affect digestibility of P (Lei et al., 1994; Lantzsch et al., 1995). There is, however, limited information about the mechanisms that regulate Ca digestibility and the role of phytate in the digestibility of Ca is not well understood.
SOURCES OF DIETARY CALCIUM
Most of the Ca in swine diets originates from inorganic sources because the concentration of Ca in cereal grains is low. However, there is limited information about the digestibility of Ca in both organic and inorganic sources of Ca when fed to pigs. In a typical corn-soybean meal diet for a 40 kg pig, the Ca contribution from corn and soybean meal is around 1 gram per kilogram of diet, whereas, approximately 5 gram Ca per kilogram diet is supplied by limestone and calcium phosphates (National Research Council, 1998).
Inorganic sources of Ca contain between 16.9 and 38.5% Ca (Table 1) and plant based feed ingredients contain between 0.01 and 0.69% Ca (Table 2). Feed ingredients of animal origin contain more Ca than plant-based feed ingredients, but usually less than the inorganic sources of Ca (Table 3). Ground limestone, calcium carbonate, and calcium phosphates are the most common Ca supplements in swine diets. If calcium carbonate is used as the standard, the relative bioavailability of Ca in limestone, aragonite, gypsum, marble dust, and oyster shell is between 93 and 102%, and in dolomitic limestone, the relative bioavailability is between 51 and 78% (Table 4; Ross et al., 1984). The low relative bioavailability of Ca in dolomitic limestone may be due to the high concentration of Mg, which may reduce the absorption of Ca because Ca and Mg use a common transport mechanism (Ross et al., 1984). Particle size does not influence the relative bioavailability of Ca in these ingredients (Ross et al., 1984).
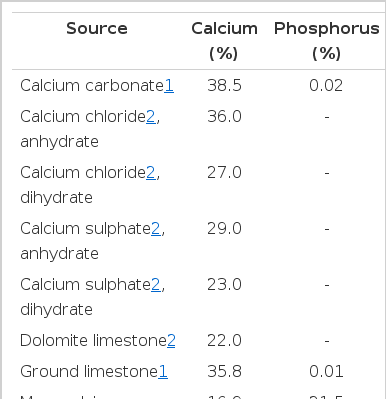
Concentration of calcium and phosphorus in inorganic sources of calcium
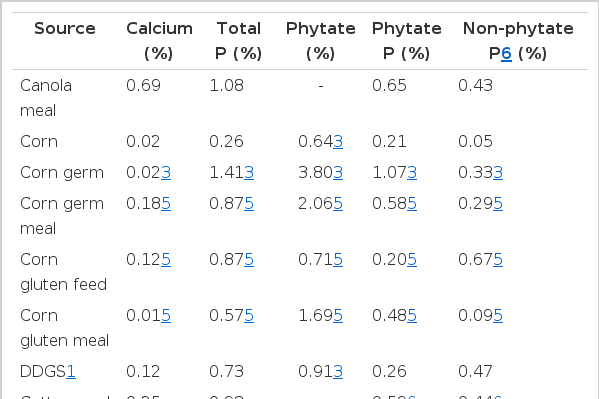
Calcium, phosphorus (P), and phytate concentration in common feed ingredients of plant origin
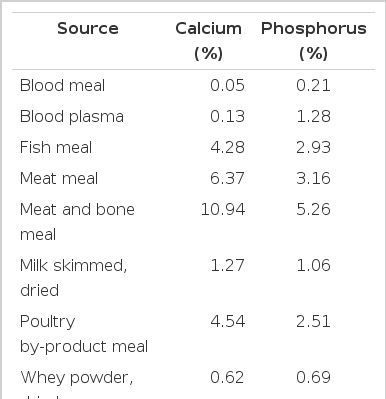

Availability and digestibility of calcium
Relative bioavailability is a comparative measure and does not always reflect the real percentage of Ca that is absorbed by the animal. Absorption of Ca may also vary with age of the animal (Hansard et al., 1961), but that is not always the case (Baker, 2011). Therefore, nutrient digestibility may be measured to more accurately estimate the availability of nutrients fed to pigs (Stein et al., 2007, 2011). The advantage of determining nutrient digestibility rather than the relative bio-availability is that the proportion of the nutrient that is absorbed by the animal is determined, and the quantity of the nutrient that is excreted from the animal may also be estimated.
DIGESTIBILITY OF CALCIUM
Digestibility of nutrients is an estimate of availability if all the nutrients that disappear from the intestinal tract are absorbed (Stein et al., 2007) and if all the absorbed nutrients are available for utilization by the animal (Ammerman, 1995). Digestibility of nutrients can be defined as apparent, standardized, or true digestibility (Stein et al., 2007). To calculate standardized or true digestibility, it is necessary to measure endogenous losses, but that is not needed if values for apparent digestibility are calculated. A portion of the endogenous loss is excreted in response to the presence of DM in the intestinal tract and is independent of the diet. This portion is known as the basal endogenous loss (Stein et al., 2007). However, another portion of the endogenous loss is excreted in response to characteristics in the diet or feed ingredient and is known as the specific endogenous loss. Both basal and specific endogenous losses contribute to the total endogenous losses and are used to calculate the true digestibility. However, basal endogenous losses are sometimes easier to measure than total endogenous losses, and basal endogenous losses are used to calculate standardized digestibility (Stein et al., 2007; Almeida and Stein, 2010). One of the disadvantages of using values for apparent digestibility is that these values are not always additive in mixed diets, but for standardized digestibility, values are additive and can be used in formulation of mixed diets (Stein et al., 2005).
Digestibility can be expressed as duodenal, ileal or total tract digestibility. The digestibility of amino acids (AA) is expressed as ileal digestibility (Stein et al., 2007) and the digestibility of P may be expressed as ileal digestibility or as total tract digestibility because there is no net absorption or secretion of P in the lower gut (Bohlke et al., 2005). However, for practical reasons, total tract digestibility of P is usually determined, and values for the apparent total tract digestibility (ATTD) and standardized total tract digestibility (STTD) of P have been determined in several ingredients (Almeida and Stein, 2010, 2012; National Research Council, 2012; Rojas and Stein, 2012; Sulabo and Stein, 2013).
Endogenous losses of minerals originate from saliva (Tryon and Bibby, 1966), cells, and pancreatic and bile secretions that enter the digestive tract (Vitti and Da Silva Filho, 2010). Several methods have been used to measure endogenous losses of P in pigs (Fernández, 1995; Fan et al., 2001; Petersen and Stein, 2006; Bünzen et al., 2008). Endogenous losses of Ca in cattle (Visek et al., 1953; Hansard et al., 1957; Martz et al., 1999) and in chickens (Cowieson et al., 2004; Liu et al., 2012) have been reported. Data for total endogenous losses of Ca in pigs have also been reported (Besançon and Guéguen, 1969; Fernández; 1995; González-Vega et al., 2013), but to our knowledge, basal endogenous losses of Ca in pigs have not been reported.
Basal endogenous losses of P have been measured using a P-free diet and an average value of approximately 200 mg/kg of dry matter intake has been reported in growing pigs (Petersen and Stein, 2006; Stein et al., 2006; Widmer et al., 2007; Almeida and Stein, 2010). A specific nutrient-free diet may be used to measure basal endogenous losses of P or AA (Petersen and Stein, 2006; Stein et al., 2007), but this approach has not been used to estimate the basal endogenous losses of Ca in pigs. If there is a linear relationship between graded levels of a nutrient in the diet and the nutrient output in feces, total endogenous losses of the nutrient can be calculated as the y-intercept of the linear regression after extrapolation back to zero input of the nutrient, and true digestibility of the nutrient is represented by the slope of the regression line (Fan et al., 2001; Kil et al., 2010).
The STTD of P has been reported for some feed ingredients (Petersen and Stein, 2006; Stein et al., 2006; Widmer et al., 2007; Almeida and Stein, 2010, 2012), but for Ca, no estimates of STTD are available. The ATTD of Ca in corn, soybean meal, canola meal, meat and bone meal, and calcium carbonate (Bohlke et al., 2005; Stein et al., 2011; González-Vega et al., 2013; Sulabo and Stein, 2013) and in some diets (Kemme et al., 1997; Stein et al., 2006, 2008; Malde et al., 2010) has been reported (Table 4). To be able to convert these values to STTD or true total tract digestibility (TTTD) of Ca, there is a need for determining basal or total endogenous losses of Ca. Total endogenous losses of Ca were reported recently by González-Vega et al. (2013) using the regression procedure with canola meal diets without or with microbial phytase and it was concluded that microbial phytase does not affect total endogenous losses of Ca. Variation in values for endogenous losses have been observed among different techniques (Dilger and Adeola, 2006; Almeida and Stein, 2010) and few data has been reported for endogenous losses of Ca. Values of 0.55 g/d (Fernández, 1995) and 1.49 g/d (Besançon and Guéguen, 1969) of total endogenous losses of Ca were obtained using isotope dilution techniques, and values of 0.11 to 0.13 g/d of total endogenous losses of Ca were reported using the regression procedure (González-Vega et al., 2013).
The calcium-phytate complex
The majority of the P in plant ingredients is bound to phytate, and 1/3 of dietary Ca may also be bound to this molecule, which results in a limited availability of both minerals (Selle et al., 2009). The presence of phytate in typical swine diets is often between 7 and 10 g/kg of diet (Selle et al., 2009; National Research Council, 2012); therefore, phytate significantly affects the digestibility of Ca and P. The negative effect of extra Ca in the diet on the efficacy of phytase (Lei et al., 1994; Lantzsch et al., 1995; Brady et al., 2002; Selle et al., 2009) may be explained by 3 mechanisms (Liu et al., 1998). One possible mechanism is the formation of insoluble Ca-phytate complexes in the small intestine (Wise, 1983; Fisher, 1992; Selle et al., 2009). Another possible mechanism is that high concentration of Ca increases pH in the intestinal content, which reduces the efficacy of phytase (Sandberg et al., 1993; Selle et al., 2009). The third possible mechanism is that extra Ca in the diet may compete for the active site of phytase and thereby reduce the efficacy of phytase in hydrolyzing phytate (Wise, 1983; Pointillart et al., 1985; Qian et al., 1996). More research is needed to elucidate the interactions among Ca, phytate, and phytase and to determine the amount of Ca that is needed in the diets.
ABSORPTION OF CALCIUM
Most dietary Ca is absorbed in the small intestine (Partridge, 1978; Liu et al., 2000), but results of some studies have indicated that Ca may also be absorbed in the colon (Liu et al., 2000). The specific region in the small intestine where Ca is absorbed is not well defined because although the absorption of Ca is greater in the proximal fourth of the small intestine than in the remaining part (Moore and Tyler, 1955a; b), the type of diet that is fed may affect the location in the intestine where Ca is absorbed (Partridge, 1978).
Before Ca is absorbed, Ca bound to other dietary compounds needs to be released in a soluble or ionic form. Gastric acids and pH-dependent enzymes assist in releasing this Ca. The pH of duodenum and jejunum is lower than in ileum of weaned pigs, 5.7, 5.9, and 6.9, respectively (Li et al., 2008). Calcium-binding proteins are responsible for the transport of Ca through the enterocyte (Bronner, 1998) and are present in greater concentrations in the duodenum than in the jejunum (Allen, 1982). Therefore, the pH and the presence of Ca-binding proteins in the enterocytes of the duodenum and jejunum may explain the greater efficacy of Ca absorption in the proximal small intestine (Allen, 1982).
Calcium is absorbed in the intestine by both nonsaturable paracellular (diffusion) and saturable transcellular (active transport) mechanisms (Bronner, 1987). The saturable mechanism occurs mainly in the proximal small intestine (duodenum and upper jejunum; Bronner, 1987). This mechanism is influenced by the amount of Ca in the diet and is most active when Ca intake is low relative to the requirement (Bronner, 1987). Paracellular absorption occurs along the small intestine between the mucosal cells, and although the nonsaturable mechanism is independent of the diet, the amount of Ca moved by this mechanism will be greater if Ca intake is relatively high compared with the requirement (Bronner, 1987).
The saturable transcellular mechanism in the small intestine absorbs Ca following 3 steps (Bronner, 1998; R & D Systems Inc., 2007). First, Ca in the lumen enters the enterocyte by channel proteins that are located in the membrane. These channels are open if the luminal Ca concentration is low and the number of channels increase in response to vitamin D. Calcium is then transported through the enterocyte by Ca-binding proteins such as calbindins, which are regulated by vitamin D. The Ca transported by the calbindins is delivered to a Ca-Na exchanger, which exchanges 3 ions of extracellular Na for 1 ion of Ca, or to a Ca-ATPase, which pumps Ca out of the cell. Both the Ca-Na exchanger and the Ca-ATPase are located on the basolateral membrane of the enterocyte. Vitamin D plays an important role in the expression of genes for the channel proteins and also in the expression of the Ca-ATPase (Bronner, 1998; R & D Systems Inc., 2007).
Increasing the level of Ca in the diet (Liu et al., 1998) and inclusion of phytase in swine diets (Lantzsch et al., 1995; Almeida and Stein, 2010; Poulsen et al., 2010) increase apparent Ca absorption. Increasing Ca levels relative to the requirement does not affect the ATTD of Ca (Stein et al., 2011), but ATTD of Ca is affected by Ca level if the levels of Ca are lower than the requirement because the endogenous losses of Ca represents a greater proportion of the Ca output (González-Vega et al., 2013). Absorption of Ca may be reduced by oxalates that may bind Ca (Weaver et al., 1987). Sulfates may also reduce Ca absorption, which may be important to consider if ingredients such as corn distillers dried grains with solubles, that may be high in S, are included in the diet (Pineda et al., 2008). Magnesium may directly reduce Ca absorption because Ca and Mg use a common transport mechanism, which explains the low bio-availability of Ca in dolomitic limestone (Ross et al., 1984). Also, high levels of Sr in the diets may reduce the absorption of Ca in broiler chicks (Corradino et al., 1971). Calcium absorption may be increased if antibiotics are supplied in diets fed to chickens (Migicovsky et al., 1951) and to rats (Heggeness, 1959). Likewise, some fibers (Weaver et al., 2010), some AA (Wasserman et al., 1956), and prebiotics increase absorption of Ca (Scholz-Ahrens et al., 2007).
METABOLISM OF CALCIUM
Between 30 and 70% of the Ca in the diet is usually absorbed, and between 65 to 95% of this Ca is retained by the pig (Kornegay, 1985; Stein et al., 2011). Retained Ca is mainly used for development and maintenance of the skeleton. However, small amounts of Ca are also used for blood clotting, enzyme activation, muscle contraction, and other functions (Soares, 1995; Taylor and Bushinsky, 2009). After Ca is absorbed, it circulates in body fluids. Approximately 50% of the Ca in serum is free as ionized Ca, 45% is bound to albumin or globulins, and the remaining 5% is present in other forms (Bringhurst and Leder, 2006). The percentage of Ca that circulates through the bones and is deposited in the bone tissue depends on the level of Ca in the diet (Bronner and Stein, 1992). Likewise, the percentage of Ca that circulates in kidney that is either recirculated or excreted in the urine is influenced by the level of Ca and P in the diet (Stein et al., 2006, 2011; Vitti et al., 2010).
Bone
Bone is composed of organic and inorganic components. Approximately one third of the weight of bone consists of organic material such as collagen and glycosaminoglycans. This fraction is important for maintaining the structure of bone because these components provide resilience to the bones (Frandson et al., 2009). The other two thirds of bone weight is composed of inorganic material, such as Ca and P salts (Frandson et al., 2009). The Ca:P ratio in bones is approximately 2.2:1 (Crenshaw, 2001). Eighty percent of this inorganic fraction consists of calcium phosphate, which is mainly present as hydroxyapatite crystals (Ca10[PO4]6[OH]2), and the remaining 20% is mainly calcium carbonate and magnesium phosphate (Frandson et al., 2009). The inorganic fraction provides rigidity to the bones (Frandson et al., 2009). Deposition of Ca and P in bones is interdependent, which means that both minerals have to be present for deposition to occur (Crenshaw, 2001).
Bone tissue is continuously degraded and synthesized and osteoclasts, osteoblasts, and osteocytes are involved in this process. Osteoblasts are bone cells responsible for synthesis of the bones, osteoclasts are bone cells responsible for resorption of the bones, and osteocytes are mature cells that are derived from the osteoblasts (Frandson et al., 2009; Veum, 2010). Osteocytes are involved in the deposition and resorption of bone tissue, and are also responsible for the transduction of signals involved in these processes (Burger et al., 1995). The activity of these cells is regulated by parathyroid hormone (PTH) and calcitonin, which are secreted by the parathyroid gland and by the C cells of the thyroid gland, respectively, depending on the concentration of Ca in plasma (Frandson et al., 2009; Sherwood et al., 2013). When concentrations of Ca in plasma are lower than normal, PTH is secreted, which increases the resorption of Ca from bones by increasing the activity of osteoclasts and inhibiting the activity of osteoblasts. Parathyroid hormone also acts in the kidneys by stimulating Ca recycling and inhibiting phosphate recycling. The enzyme 1α-hydroxylase, which activates vitamin D to 1,25-dihydroxycholecalciferol, is also stimulated, which results in increased Ca absorption from the intestines (Costanzo, 2006), because 1,25-dihydroxycholecalciferol may enhance the synthesis of calcium binding proteins (Kumar, 1995) and calcium channels in the enterocyte (Bouillon et al., 2003).
If concentrations of Ca in plasma are greater than normal, calcitonin is secreted, which inhibits osteoclastic activity and reduces Ca release from bone (Costanzo, 2006). Excretion of Ca via the kidneys is also increased, and the uptake of Ca by cells is increased (Crenshaw, 2001).
Renal Ca metabolism
The kidneys play an important role in the regulation of Ca by filtering, reabsorbing, and excreting the Ca in blood to maintain stable Ca levels in plasma (Taylor and Bushinsky, 2009). Calcium is absorbed regardless of the dietary Ca and P concentration, but if Ca is absorbed in excess of the requirement, excesses are excreted in the urine (Stein et al., 2006, 2011). Pigs fed a P-free diet were able to absorb Ca in the intestines, but because there was not enough P to retain the absorbed Ca in bones, the absorbed Ca was excreted in the urine (Stein et al., 2006). This indicates that Ca homeostasis is not regulated in the intestines; instead, the kidneys appear to be the main regulatory organs. The Ca:P ratio plays an important role in urine excretion of these minerals. If the Ca:P ratio is greater than that needed for bone tissue synthesis, Ca excretion in the urine is increased due to the low availability of P (Helander et al., 1996; Stein et al., 2006). However, if diets with a low Ca:P ratio are fed, most of the P is excreted in the urine due to a lack of Ca for bone tissue synthesis (Stein et al., 2011). Although the body can regulate the levels of Ca in plasma, prolonged consumption of diets with excess Ca or deficiency of Ca can cause serious problems such as kidney stones or bone diseases (Taylor and Bushinsky, 2009).
Not all the Ca in the blood is recycled by the kidneys because Ca that is bound to albumin is not filtered by the glomerulus (R & D Systems Inc., 2007). If Ca is filtered by the glomerulus, approximately 60% of the Ca is resorbed in the proximal tubule, 10 to 15% in the ascending loop of Henle, and the remaining 15 to 25% in the distal convoluted tubule and collecting duct (Kumar, 1995).
CONCLUSION
Calcium is needed for bone and teeth formation and for many other physiological functions in the body. Plasma Ca levels are mainly regulated by PTH and calcitonin that have major effects on Ca metabolism in intestines, bones, and kidneys. For the retention of Ca and P in the body, both minerals need to be available in bone cells, and a low concentration of one mineral will prevent bone synthesis and cause excretion of the other mineral in the urine. Most of the Ca needs of pigs fed commercial diets is usually supplemented by limestone, calcium carbonate, and calcium phosphates because of the low concentration of Ca in most feed ingredients of plant origin. The supplemented Ca can be from an organic or an inorganic source, and to formulate diets, it is important to consider the digestibility of Ca in these sources. More information about Ca digestibility in ingredients included in diets fed to pigs is needed to improve the utilization of both Ca and P by the animals. Because most ingredients from plant origin contain phytic acid, the effect of phytic acid on Ca digestibility needs to be clarified. However, to determine if apparent, standardized, or true digestibility values are most accurate, endogenous losses of Ca from the gastrointestinal tract of the pig need to be determined, and additivity of digestibility values from individual ingredients used in mixed diets need to be confirmed. To determine if ileal or total tract digestibility values most accurately describe the absorption of Ca, the site of absorption also needs to be described.
Notes
A part of this manuscript was presented in the M.S. thesis of J. C. González-Vega (2012).