Effects of Immune Stress on Performance Parameters, Intestinal Enzyme Activity and mRNA Expression of Intestinal Transporters in Broiler Chickens
Article information
Abstract
Immune stress is the loss of immune homeostasis caused by external forces. The purpose of this experiment was to investigate the effects of immune stress on the growth performance, small intestinal enzymes and peristalsis rate, and mRNA expression of nutrient transporters in broiler chickens. Four hundred and thirty-two 1-d-old broilers (Cobb500) were randomly assigned to four groups for treatment; each group included nine cages with 12 birds per cage. Group 1 = no vaccine (NV); Group 2 = conventional vaccine (CV); group 3 = lipopolysaccharide (LPS)+conventional vaccine (LPS); group 4 = cyclophosphamide (CYP)+conventional vaccine (CYP). The results demonstrated that immune stress by LPS and CYP reduced body weight gain (BWG), feed intake (FI), small intestine peristalsis rate and sIgA content in small intestinal digesta (p<0.05). However, the feed conversion ratio (FCR) remained unchanged during the feeding period. LPS and CYP increased intestinal enzyme activity, relative expression of SGLT-1, CaBP-D28k and L-FABP mRNAs (p<0.05). LPS and CYP injection had a negative effect on the growth performance of healthy broiler chickens. The present study demonstrated that NV and CV could improve growth performance while enzyme activity in small intestine and relative expression of nutrient transporter mRNA of NV and CV were decreased in the conditions of a controlled rational feeding environment. It is generally recommended that broilers only need to be vaccinated for the diseases to which they might be exposed.
INTRODUCTION
It is apparent that many factors can influence the immune response of an animal under stress. Stress can suppress, enhance or have no effect on the immune response of an animal. There are many interacting factors influencing the immunological response of an animal under stress, these include stressor types (psychological vs physiological vs physical), duration of stressor (chronic vs acute), genetics, age, and immune status. Some factors in the homeostasis of broilers directly or indirectly affect their immune status, and immunosuppression induced by stress is manifested by failures in vaccination and increased morbidity and mortality of flocks (Shini et al., 2010). Vaccinations are considered necessary to reduce the mortality rate that occurs in modern poultry production. However, immunizations have sometimes been administered in large doses at a high frequency and improper vaccination may lead to immune stress. Studies have demonstrated that stress could affect the intestinal function in animals (Apajalathi and Kettunen, 2003; Teo and Tan, 2007; Rajkumar et al., 2011; Wilkinson et al., 2011) and furthermore disturb the absorption of nutrients (Garriga et al., 2006; Jacobi et al., 2006; Albin et al., 2007). The activities of digestive enzymes are such important factors affecting the absorption and utilization of nutrients in the alimentary tract that they are often used to indicate the intestine function of nutrient absorption. Heat stress causes metabolic and physiological changes in the intestinal transport of glucose through the increase of SGLT1 (Carriga et al., 2006). The effects of stress on the intestinal nutrient absorption in animals may be reflected in two ways. One is nonspecific, in which the absorption of nutrients is affected by the changes of digestive enzymes and intestinal peristalsis rate (Tsukada et al., 2002); the other is specific, in which the absorption of nutrients is affected by changing the expression of transporters particularly of nutrients (Shepherd et al., 2004). Expression of these proteins varies during the development of chickens at different ages (Gilbert et al., 2007; Duarte et al., 2011; Yadgary et al., 2011). Research has shown that the immune response can affect growth performance and enteric diseases in commercial poultry, contribute to losses in productivity, increase mortality, and increase the contamination of products for human consumption (Yang et al., 2011). Previous studies have yielded various results; however, the mechanisms of these phenomena have not yet been fully understood. To our knowledge, few studies on the potential effect on the status of broilers reared under immune stress conditions were available. The aims of this study were to determine the balance between the hypo- and hyper-stress, minimize distress while increasing conditions for eustress.
MATERIALS AND METHODS
Animals and experimental design
The experiment was performed as a single-factor trial. Four hundred thirty-two 1-d-old broilers (Cobb500) were randomly assigned to four treatments with nine pens per treatment and twelve birds per pen. The four treatments were control group with no vaccine (NV), conventional vaccine group (CV), lipopolysaccharide injected+ conventional immunization group (LPS), and cyclophosphamide injected+conventional immunization group (CYP). All birds were housed in an environmentally controlled room with an initial room temperature of 34°C for the first 5 d and gradually decreased to 21°C by the end of the experiment at the third week. Birds were provided with feed and water ad libitum. The lighting program was 24 h light for the first day, followed by 18 h light/6 h dark for the rest of the trial period. All experimental procedures were approved by the Animal Ethics Committee of the Northwest A & F University.
Chemicals and vaccines
The chemicals used in this study included LPS from E. coli (L2880, Sigma-Aldrich, Inc., St. Louis, Missouri, USA) and CYP (C0768, Sigma-Aldrich). The vaccines used in this study included infectious bronchitis vaccine (IBH 120), the inactivated avian influenza (AI) vaccine (H5N2, N28 strain), the live vaccine strain Clone-30 of the Newcastle disease virus (NDV), the combined inactive vaccines for infectious bursal disease and the Newcastle disease (ND-IB), all of them were supplied by Harbin Veterinary Research Institute (Harbin, China).
Growth performance measurement
Body weight gain (BWG) and feed intake (FI) were recorded based on cage weekly. Mortality was recorded daily and feed conversion ratio (FCR) was corrected according to mortality.
Sample collection
Six birds with body weight close to the mean from each replicate were killed by cervical dislocation on d 21 and d 42. The duodenum, jejunum and ileum were weighed. Digesta from the duodenum, jejunum and ileum was put in ice-cold phosphate buffered saline (PBS, pH 7.4). Samples were centrifuged (13,000×g for 5 min) at 4°C, and supernatant was frozen in a 2 ml eppendorf tube immediately at −20°C until needed. The activities of amylase, lipase and trypsin in the small intestinal digesta were determined using standard kits (Jiancheng Bioengineering Institute, Nanjing, China). Intestinal peristalsis rate was detected according to the method described by Poggioli et al. (1995). The level of digesta sIgA was examined with the chicken IgA ELISA quantitation kit (no. E30–103, Behyl Laboratories Inc., Montgomery, TX, USA) as mentioned by Schuijffel et al. (2005).
RNA extraction and real-time PCR (RT-PCR)
Total RNA was extracted from individual intestinal tissue (duodenum, jejunum and ileum) using the RNeasy kit (Kim et al., 2010) (Qiagen, Germantown, MD, USA). The samples from small intestine were removed and deposited in a −80°C freezer for further analyses of SGLT1, CaBP-D28k and L-FABP. A 20 to 30 mg aliquot of each sample was weighed and kept on dry ice until homogenization according to manufacturer’s recommendation. In the extraction, RNA was eluted by rinsing the column membrane twice with 50 μl of RNase-free water. Total RNA concentration was determined by optical density (OD) 260 (NanoDrop-1000, Thermo Fisher Scientific, Waltham, MA, USA), and RNA purity was verified by OD260/OD280 value and formaldehyde-agarose gel electrophoresis. The first strand cDNA was stored at −20°C. The relative abundance of transporter mRNA was assayed by quantitative RT-PCR (Table 1).

Primer sequences for PCR amplification of target genes
Gene expression of SGLT-1, CaBP-D28k and L-FABP was assessed by 2-step quantitative RT-PCR using the SYBR Green PCR master mix (Applied Bio Systems Inc., Foster City, CA); mRNA expression was quantified using the iCycler iQTM5 multicolor RT-PCR detection system (Bio-Rad Laboratories, Hercules, CA, USA). A 25 μl PCR mixture was set up as follows: cDNA, 1 μl; each primer (10 pmol/ml), 1 μl; 2×SYBR Green quantitative PCR master mix, 12.5 μl; and double-distilled water, 10.5 μl. The temperature cycles were set as following: 95°C for 30 s, followed by 40 cycles at 95°C for 5 s and 60°C for 30 s. SYBR green fluorescence was measured at the end of each cycle. The melting curve was recorded at 60°C. All primer concentrations were optimized before actual runs. All measurements were carried out in triplicate to obtain the average values. Expression of SGLT-1, CaBP-D28k, and L-FABP mRNAs from different parts of small intestine was calculated by the 2−ΔΔCt method (Livak and Schmittgen, 2001).
Statistical analysis
Means of replicate were used as the experimental unit for statistical analysis. Data were subjected to one-way ANOVA analysis for the differences between the four groups using SAS software (SAS Institute Inc., Cary, North Carolina, USA, 2002). Differences between means at p< 0.05 were considered to be statistically significant.
RESULTS
Performance parameters
The effect of different types of immune stress on feed intake (FI), body weight gains (BWG) and the feed conversion ratio (FCR) is presented in Table 2. No differences were observed in FI and BWG between NV group and CV group (p>0.05). However, compared with NV, the BWG decreased significantly in the chickens that had been treated with LPS or CYP (p<0.05 and p<0.01), and the FI decreased significantly in the chickens that had been treated with LPS. The FCR showed no significant difference among the four groups (p>0.05).
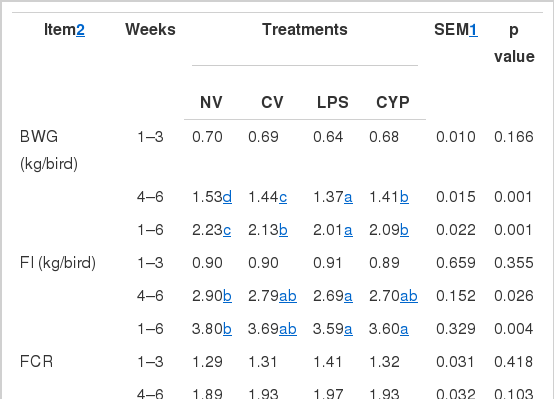
Measurements of organ relative weight, enteric enzyme and intestinal peristaltic rate
Relative weight indices of duodenum and jejunum in NV group were higher than that in LPS group on d 21 (p<0.05). There was no significant difference in the duodenum, jejunum and ileum weight indices on d 42 (Table 3).
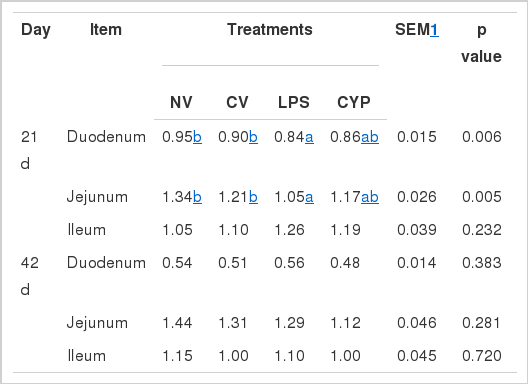
Effects of immunity stress on small intestinal index in broilers (mg/g)
As shown in Table 4, the amylase and lipase of jejunum and ileum in CYP treatment were higher compared with the NV and CV treatments on d 21 and 42 (p<0.05 and p<0.01), while the trypsin activity in the small intestine was not significantly different during the whole period (p>0.05). The intestinal peristalsis rate and sIgA content in NV group were significantly higher than those in LPS and CYP groups (p<0.01) (Table 5).
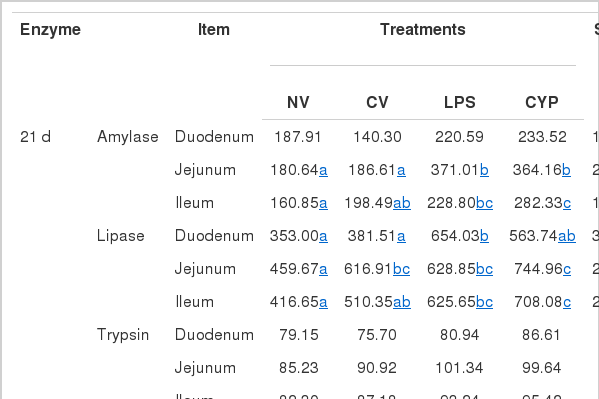
Effects of immunity stress on digestive enzyme activity in small intestinal digesta (U/mg)
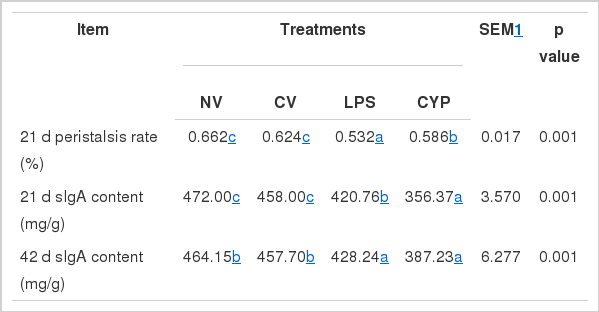
Effects of immunity stress on intestinal motility and digesta sIgA in broilers
Effects of stress on the expression of SGLT1, CaBP-D28k and L-FABP mRNAs in small intestine
The mRNA levels of related genes in the small intestine are shown in Table 6. The relative expression of SGLT-1, CaBP-D28k and L-FABP mRNAs in the three parts of small intestine in the LPS and CYP groups was significantly or numerically up-regulated in comparison to CV and NV groups (p<0.05).

Effects of immunity stress on the relative expression of SGLT1, CaBP-D28k and L-FABP mRNAs in the different parts of intestine
DISSCUSSION
Our results showed that immune stress decreased the FI and BWG of chickens (Table 2). It was reported that immune stress may decrease the growth performance of broilers (Eid et al., 2003; Lin et al., 2004; Virden et al., 2007; Star et al., 2008; Rajapakse et al., 2010). The depressed growth performance might be explained by which immune stress is the immuno-modulator and allocates nutrients toward immune response.
Dudley et al. (1998) observed that dietary restriction leads to a decrease in intestinal protein synthesis with concomitant lowering of intestinal weight. Thermal stress caused a consistent increase in amylase activity in broilers, which could result in a significant increase in starch hydrolysis (Osman et al., 1983). Other studies, however, reported that thermal stressed birds had a higher enzymatic activity (Routman et al., 2003). Furthermore, compared to birds fed with control diets, birds with early feed restriction and exogenous protease and amylase supplementation have increased maltase, lipase, and trypsin immediately after the restriction period (Pinheiro et al., 2004). In fact, birds subjected to feed restriction will increase intestinal absorption, and the gastrointestinal tract will be spared the effect of restriction on weight that affects other organs and the BWG (Pinheiro et al., 2004). These responses depend on the age of animal and the duration of the restriction period (Casirola et al., 1997; Ferraris et al., 2001). These results may indicate the adaptation of the enzymatic response to immune stress, the higher enzyme activity may have been stimulated by the presence of feed in the gastrointestinal tract for a longer time due to a slower rate of feed passage through the gastrointestinal tract (Barash et al., 1992). Moreover, some investigators e.g. Lessire et al. (1990) observed that feed restriction tended to improve protein, fat, and starch digestibilities irrespective of the diet. However, slower movement speed can increase the chyme residence time in the gut and help nutrient absorption. The current research demonstrated that the effect of immune stress on chickens increased the stress stimulation in the intestinal tract, which might lead to the increased production of intestinal amylase and lipase enzymes in the LPS and CYP groups.
The intestine is relatively enriched with cells actively secreting IgA, and sIgAs from the lamina propria of villi in the duodenum and jejunum, and Peyer’s patches are important components of intestinal mucosal immunity (Mestecky et al., 1987). Mucosal immunity is an important part of the humoral immunity. Secretory IgA is the most prominent antibody present at mucosal surfaces, and is the factor of mucosal immunity that provides passive immunoprotection against invading pathogens in the gastrointestinal tract. The results of this study showed that injection with LPS and CYP could decrease sIgA content compared with the NV group. This implies that LPS and CYP may impair the humoral immune system, which led to small intestinal mucosal lesions.
Nutrient transporters play a critical role in the absorption of luminal substrate. In the small intestine of animals, SGLT1 is the key factor that affects absorption of glucose in the small intestine (Kellett, 2001; Wood and Frayhum, 2003). A previous study showed that glucocorticoids released during stress periods induce the expression of glucocorticoid regulated kinase, which enhances glucose transportation by increasing SGLT1 abundance in the cell membrane (Carriga et al., 2006). Gilbert et al. (2007) also observed an increase in mRNA abundance of SGLT1 in feed restricted chickens. SGLT1 has the unique ability to transport glucose against its concentration gradient, and it is considered the major pathway for glucose assimilation in the small intestine. Thus, a lower luminal sugar concentration than that in blood during feed restriction may cause the increase in this transporter, due to its important role in the assimilation of glucose. Calcium is absorbed in the duodenum and jejunum by calcium-binding protein (CaBP), which is regulated by a 28 KDa 1, 25(OH)-D3 protein (CaBP-D28k) (Craviso et al., 1987). Two types of fatty acid binding protein (FABP) are found in intestinal epithelium, i. e. intestinal FABP (I-FABP) and liver FABP (L-FABP) (Banaszak et al., 1994). I-FABP participates in the uptake of long-chain fatty acids from digesta in the small intestine into intestinal epithelial cells, and L-FABP transports fatty acids from the cells to the tissues (Prows et al., 1995). In the present study, the increasing demand for energy causes changes in the intestinal epithelium and gene expression of nutrient transporters. The upregulated expression of CaBP-D28k and L-FABP genes in stressed birds may be related to an increased intestinal absorptive function due to immune stress. The increased expression of nutrient transporter mRNA during stress is likely to be a compensation for the loss of absorptive area in the small intestine.
It can be concluded that broilers with NV or CV can achieve higher growth performance in a controlled, rational feeding environment, while immune stress can interrupt the homeostasis of the intestine and impair intestinal mucosal immune function. However increased enzyme and mRNA expression in chickens under stress might compensate for the nutrient consumption by the system immune responses or basic metabolic requirements. Extra vaccinating will cost extra growth. This study provides some details about the relationship between immune stress and nutrient partition.
ACKNOWLEDGEMENTS
The authors are grateful to the support from the Science & Technological Projects of Shaanxi Province, China (2010ZDGC-02, 2011KTCQ02-02), and National Natural Science Foundation of China (No.31001017 and No.31172223).