Molecular Characterization and Expression Analysis of the Peroxisome Proliferator Activated Receptor Delta (PPARδ) Gene before and after Exercise in Horse
Article information
Abstract
While athletic abilities such as speed, endurance and recovery are important in the horse, genes related to these abilities have not been extensively investigated. Here, we characterized the horse peroxisome proliferator-activated receptor delta (PPARδ) gene and analyzed the expression of PPARδ during exercise. PPARδ is a known regulator of β-oxidation, muscle fiber transformation, and running endurance. Through evolutionary analysis using the synonymous and non-synonymous mutation ratio, it was revealed that positive selection occurred in the horse PPARδ gene. Two important domains related to nuclear hormone receptors, C4 zinc finger and ligand binding domain, were also found to be conserved well in horse PPARδ. Horse PPARδ was expressed ubiquitously in many tissues, but the expression level was various depending on the tissues. In the skeletal muscle, PPARδ increased about 2.5 folds after 30 min of exercise. Unlike in muscle, the increase of PPARδ expression was observed at 60 min but not 30 min of exercise in leukocytes. This finding might be useful for testing the endurance of horse using blood samples. Conclusively, the horse PPARδ gene is evolutionarily conserved well and can be used as a biomarker of endurance in horse.
INTRODUCTION
Many economic traits including speed, robustness, and endurance of racing horses are related to exercise. Previously, whole transcriptome analysis using RNA-sequencing in six thoroughbred horses before and after exercise was performed to identify exercise-related genes (Park et al., 2012). Differentially expressed gene analysis of skeletal muscle and blood using RNA-sequencing of selected genes regarded as associated with economically important traits of horses has become an interest for the purposes of selection and improvement.
Peroxisome proliferator-activated receptors (PPAR) are members of the nuclear receptor superfamily and was first identified in xenopus (Krey et al., 1993). PPARs can be activated and cooperate with thyroid hormone and retinoid X receptors by the formation of a heterodimer complex for transcriptional regulation of target genes (Wahli et al., 1995). PPARs play essential roles in the regulation of cellular differentiation, development, metabolism (carbohydrate, lipid, protein), and tumorigenesis of higher organisms (Berger and Moller, 2002; Belfiore et al., 2009). Among the PPARs, PPARδ has an important role in fat metabolism in muscle; in knockout mice studies, it has been shown that it activates oxidation of fatty acid in muscle while increases conversion of glucose to fatty acid in liver (Lee et al., 2006). The overexpression of PPARδ resulted in an increased oxidative metabolism and hyperplasia causing the change of fiber composition in mouse muscle (Luquet et al., 2003). Together with its metabolic role in muscle, PPARδ acts as an important regulator of satellite cell proliferation and muscle regeneration (Angione et al., 2011).
According to previous studies, expression of PPARδ mRNA and protein is increased in the skeletal muscle after endurance exercise (Luquet et al., 2003; Mahoney et al., 2005). PPAR’s expression in skeletal muscle is related to the late induction of exercise-induced responses during endurance training, such as increased oxidative capacity of type I muscle fibers and mitochondrial biogenesis (Wang et al., 2004). The overexpression of PPARδ in mouse skeletal muscle induces the up-regulation of gene expression related to slow fiber contractile proteins, mitochondrial biogenesis, and β-oxidation. Therefore, the rapid and regulated changes of PPARδ expression could improve physical performance. Despite extensive and numerous studies on the function and expression of the PPARδ gene in various species, it has not been researched in horses. The objective of this study was to characterize the horse PPARδ gene at the molecular level by performing bioinformatics and the real-time polymerase chain reaction (RT-PCR) analysis, and to investigate the transcriptional regulation of PPARδ gene after exercise and the possibility of PPARδ as a biomarker.
MATERIALS AND METHODS
Bioinformatic analysis of horse peroxisome proliferator-activated receptor delta gene
The coding DNA and amino acid sequences of PPARδ of various species (horse, human, chimpanzee, mouse, rat, cow, pig, chicken, dog, and cat) were retrieved from Ensembl (http://www.ensembl.org/). The amino acid sequences were aligned using ClustalW in the BioEdit software. The protein domains were predicted by using the SMART domain search program (http://smart.embl-heidelberg.de/). Phylogenetic analysis data was obtained by using the Neighbor-Joining method (Saitou and Nei, 1987) in MEGA software (Tamura et al., 2011). The parameters were as follows: pairwise-deletion, 1000 bootstrap replications, and Kimura 2. Pairwise deletion was chosen to retain all such sites initially, excluding them as necessary in the pairwise distance estimation. The group within tree was used for 1000 bootstrap replications to derive the reliability value. Substitutions of nucleotides were obtained by using the Kimura 2-parameter model. We also showed synonymous substitutions per site (Ks) and non-synonymous substitutions per site (Ka) in PPARδ gene for individual variation.
Horse blood and tissue samples
Blood and muscle samples were collected from three thoroughbred horses, which were 2 to 4 year-old geldings and maintained at Ham-an Racing Horse Resort & Training Center. Samples were collected before and after 30 min and 60 min of trotting. Exercise was performed about 13 km/h of speed on a treadmill. The National Institute of Subtropical Agriculture, Rural Development Administration, Republic of Korea, provided three Jeju horses which were used for tissue sampling. Skeletal muscle, kidney, thyroid, lung, appendix, colon, spinal cord and heart tissues were kept in a liquid nitrogen tank until RNA extraction. Animal care and sample collection were conducted in accordance with protocols approved by the Institutional Animal Care and Use Committee of the Pusan National University (PNU-2014-0644 and PNU-2014-0645).
RNA extraction and cDNA synthesis
Trizol reagent (Invitrogen, Carlsbad, CA, USA) was used to extract total RNA from tissue samples and leukocytes according to the manufacturer’s manual. In order to prevent contamination of genomic DNA, all RNA samples were treated with RNase-free DNase (Qiagen, Venlo, Netherlands). Next, the RNA samples were quantified by using a NanoDrop ND-1000 Spectrophotometer (Thermo Fisher Scientific Inc., Waltham, MA, USA). The cDNA was synthesized from the each RNA sample by moloney-murine leukemia virus reverse transcriptase (Promega, Madison, WI, USA) in a reaction containing oligo-dT primers and RNase inhibitor (Promega, USA) for 4 hours incubating at 37°C.
Real time-quantitative polymerase chain reaction analysis
Firstly, PCR was conducted to verify the removal of genomic DNA and the synthesis of cDNA. The PPARδ and glyceraldehyde 3-phosphate dehydrogenase (GAPDH). primer sets for PCR were designed by using the PRIMER3 software (http://frodo.wi.mit.edu/primer3/). The sequences of PPARδ primers were 5′-AGC AAC CTC TTC CTC AAC GA-3′ for forward primer and 5′-GAG GCT GCG TAG GAA CTC AC-3′ for reverse primer. The PCR product size of PPARδ was expected to be 150 bp. The PCR conditions consisted of an initial step of 94°C for 10 min, 35 cycles of 94°C for 30 s, 60°C for 30 s, 72°C for 30 s, and a final step of 72°C for 10 min. The PCR products were analyzed by electrophoresis on a 2.0% Seakem LE agarose gel.
To quantify the PPARδ gene in horse tissues and leukocytes, RT-quantitative polymerase chain reaction (qPCR) was conducted by using CFX-96 real-time PCR (BioRad, Hercules, CA, USA). Each reaction was executed in 25 μL of a total volume containing 14 μL of SYBR green master mix, 2 μL of forward primer (5 pmol), 2 μL of reverse primer (5 pmol), 5 μL of distilled water, and 2 μL of the cDNA (50 ng/μL). The PCR conditions were as follows: a pre-denaturation step at 94°C for 3 min, 39 cycles at 94°C for 10 s, 60°C for 30 s, and 72°C for 30 s, followed by a final step at 72°C for 10 min. Dissociation was accomplished in conditions where temperature increased from 60°C to 95°C for over 25 min (linear increase of 0.5°C). All samples were measured in triplicate to ensure reproducibility, and the expression levels were calculated from Ct value by using the 2−ΔΔCt method (Livak and Schmittgen, 2001). The GAPDH gene was used for reference.
Statistical analysis of gene expression
The significance (p<0.05) of data was determined using Student’s t-test or analysis of variance method. The difference among each value was determined by Tukey’s test with alpha = 0.05. All data values are presented as mean ±standard error of mean.
RESULTS AND DISCUSSION
The horse PPARδ gene consisted of eight exons with a total length of 1,644 bp. The translation of PPARδ starts in the third exon and stops at the last exon. In comparing the nucleotide sequences in various species (human, chimpanzee, mouse, rat, cow, pig, chicken, dog, cat), the synonymous substitution (Ks) ranged from 0.363 to 1.330, whereas the non-synonymous substitution (Ka) ranged from 0.407 to 1.553 in horse (Table 1). It is of note that the lowest (0.407) and highest (1.553) differences of Ka of horse PPARδ were obtained when horse compared with cat and horse compared with mouse respectively. Human and chimpanzee are of similar sequence and/or amino acid, showing the lowest Ks value (0.018). The average Ka value was 1.056 and that of Ks was 0.914, resulting to a Ka/Ks ratio greater than 1. This indicates that non-synonymous substitutions occurred faster than synonymous substitutions. These results are based on the assumption that the protein-coding regions in the human genome undergo positive selection during evolution (Bustamante et al., 2005). Results indicate that positive selection occurred in the PPARδ genes during evolution in horses. This ratio also suggests possible evolution of the PPARδ gene in various species.
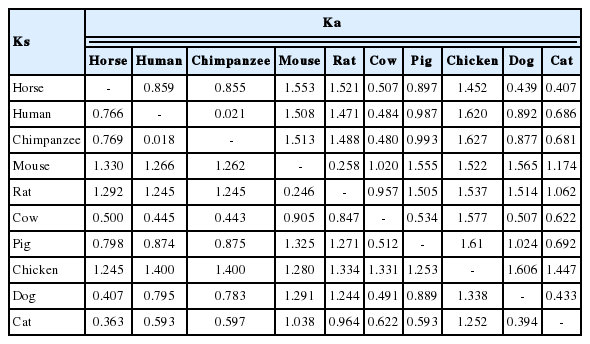
Synonymous (Ks) and non-synonymous (Ka) substitutions per site in the PPARδ gene of various species
To characterize horse PPARδ protein, amino acid sequences of PPARδ from various species were aligned and analyzed. The PPARδ amino acid sequences are highly conserved between the species except for the N-terminal of the sequence (Figure 1A). In domain search using the SMART program, it was revealed that horse PPARδ protein has two highly conserved domains, C4 zinc finger and ligand binding domains of nuclear hormone receptors. In a phylogenetic analysis with the Neighbor-Joining method, horse PPARδ is clustered in the same clade with dog and chicken while having a large genetic distance from mouse and rat (Figure 1B).

Analysis of horse PPARδ amino acid sequences. (A) Comparison of amino acid sequences of horse PPARδ in various species. In SMART domain prediction, it is revealed that C4 zinc finger (dashed box) and ligand binding (solid box) domains of nuclear hormone receptors were conserved in horse PPARδ. (B) Phylogenetic tree of PPARδ in various species. PPARδ, peroxisome proliferator-activated receptor delta.
Next, we performed RT-PCR analysis using eight different tissues from three horses to examine the gene expression pattern. All horse tissues expressed the PPARδ gene ubiquitously, and PPARδ was highly expressed in thyroid and lung while relatively low in skeletal muscle (Figure 2). It is of note that PPARδ was highly expressed in thyroid cells unlike other PPAR genes such as PPARα and PPARγ (Bookout et al., 2006). High expression of the PPARδ in thyroid and lung also corresponds to a previous expression study in xenopus (Krey et al., 1993). In human, the expression of PPARδ in primary thyroid cells was induced by serum, thyroid-stimulation hormone, and cyclic AMP (3′-5′-cyclic adenosine). PPARδ is up-regulated in human thyroid tumors and induces cell proliferation through the cyclin E1. In this sense, increased expression of PPARδ and Ki-67 is known as an in situ proliferation marker of benign and malignant human thyroid tumors (Zeng et al., 2008), where the mean expression of native PPARδ was increased by 2-fold to 5-fold and correlated with that of the in situ proliferation marker Ki67 in six different classes of benign and malignant human thyroid tumors.
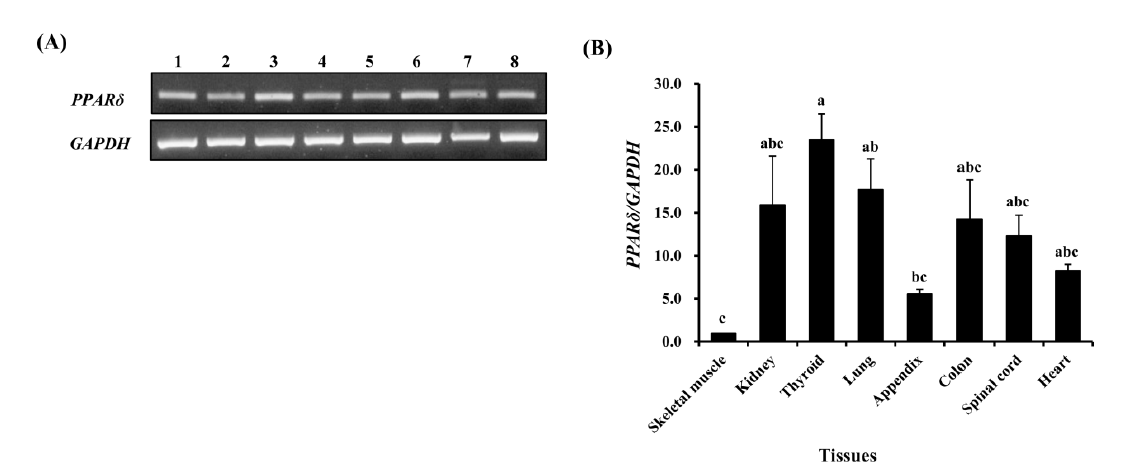
Analysis of horse PPARδ expression in various tissues of Jeju horse. (A) Expression of horse PPARδ gene in various tissues. 1: skeletal muscle, 2: kidney, 3: thyroid, 4: lung, 5: appendix, 6: colon, 7: spinal cord, and 8: heart. The bands are saturated by running the polymerase chain reaction for 35 cycles. (B) Determination of expression levels of horse PPARδ in horse tissues. Expression value was calculated using the 2−ΔΔCt method. The values are presented by mean±standard error of mean. The expression levels of the gene were various in tissues (n = 3, p<0.01). The difference of each value was determined by Tukey’s test and the bars containing same letter on the top are not significantly different among them (alpha = 0.05). GAPDH was used for normalization. PPARδ, peroxisome proliferator-activated receptor delta; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
To identify the relation between exercise and PPARδ gene expression levels, the expression pattern of the gene was analyzed in skeletal muscle and leukocytes of horse. In skeletal muscle, PPARδ was highly expressed about 2.5 fold at 30 min of exercise (Figure 3A, p<0.001). In leukocytes, expression levels of PPARδ were analyzed by RT-qPCR with the samples obtained before and after 30 min and 60 min of exercise (Figure 3B). Unlike skeletal muscle, which showed higher expression at 30 min of exercise, the expression of PPARδ increased slowly and was significantly higher at 60 min of exercise in leukocytes. It is well known that endurance exercise increases expression of PPARδ gene in the muscle of mouse and human (Luquet et al., 2003; Wang et al., 2004; Mahoney et al., 2005). This expression pattern of PPARδ was also observed in horse muscle and even in the leukocytes although it took a longer time than in muscle. The increase of PPARδ expression in horse muscle might induce the oxidative metabolism to supply the energy required for endurance exercise. As the leukocytes are relatively feasible method of collecting samples from the living horse, analysis of PPARδ expression in leukocytes may be a useful tool for indirectly scoring the muscle fatigue.
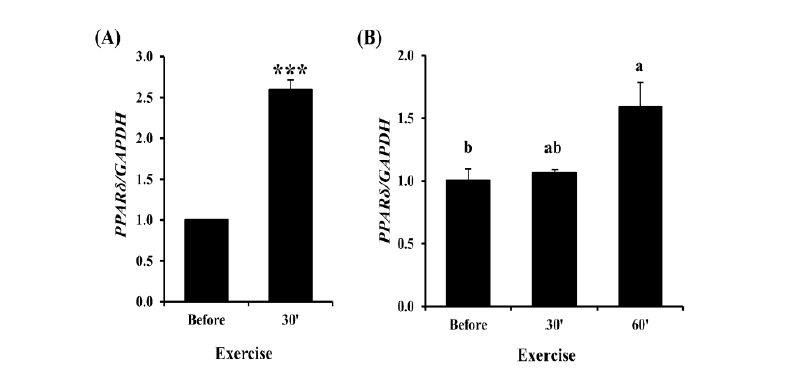
Analysis of PPARδ expression before and after exercise in skeletal muscle and leukocytes. (A) Expression level of PPARδ before and at 30 min after exercise in horse skeletal muscle. The expression of PPARδ increased about 2.5 fold at 30 min of exercise in skeletal muscle (n =5). *** means p<0.001. (B) Expression analysis of horse PPARδ before and after exercise in leukocytes. Expression value was calculated using the 2−ΔΔCt method. The values are presented by mean±standard error of mean. The expression levels of PPARδ was significantly higher after 60 min of exercise than before exercise (n = 3, p<0.05). The difference of each value was determined by Tukey’s test and the bars containing same letter on the top are not significantly different (alpha = 0.05). GAPDH was used for normalization. PPARδ, peroxisome proliferator-activated receptor delta; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
CONCLUSION
In conclusion, the horse PPARδ gene is well conserved evolutionarily and has functional domains found in other animals. The expression of PPARδ gene is regulated by endurance exercise in muscle and leukocytes, and this gene could be a useful biomarker to measure endurance and fatigue. In further studies, the analysis of gene expression related to oxidation, mitochondrial biogenesis, and enhancement of physical performance by PPARδ after exercise might be useful for understanding the relationship between PPARδ and athletic ability in horse.
ACKNOWLEDGMENTS
This work was supported by a grant from the Next-Generation BioGreen 21 Program (No. PJ011044 and PJ011173), Rural Development Administration, Republic of Korea.