Effect of Dietary Marine Microalgae (Schizochytrium) Powder on Egg Production, Blood Lipid Profiles, Egg Quality, and Fatty Acid Composition of Egg Yolk in Layers
Article information
Abstract
Two hundred and sixteen Institut de Sélection Animale (ISA) brown layers (40 wks of age) were studied for 6 wks to examine the effect of microalgae powder (MAP) on egg production, egg quality, blood lipid profile, and fatty acid concentration of egg yolk. Dietary treatments were as follows: i) CON (basal diet), ii) 0.5% MAP (CON+0.5% Schizochytrium powder), and iii) 1.0% MAP (CON+1.0% Schizochytrium powder). From 44 to 46 wks, egg production was higher in 1.0% MAP treatment than in control treatment (linear, p = 0.034); however, there was no difference on the egg production from 40 to 43 wks (p>0.05). Serum triglyceride and total cholesterol were significantly reduced in the groups fed with MAP, compared to those in groups fed with control diets (Quadratic, p = 0.034 and p = 0.039, respectively). Inclusion of 0.5% MAP in the diet of layers improved egg yolk color, compared with hens fed with basal diet at 46 wks (quadratic, p = 0.044). Eggshell thickness was linearly increased in MAP-fed treatments at 46th wk (p<0.05). Concentration of yolk docosahexaenoic acid (DHA; C22:6n-3) was increased in treatment groups fed with MAP (linear, p<0.05). The n-6 fatty acids, n-6/n-3 fatty acid, and unsaturated fatty acid/saturated fatty acid were decreased in treatment groups fed with MAP (linear, p<0.05). These results suggest that MAP improved the egg production and egg quality, and may affect serum lipid metabolites in the layers. In addition, MAP increases yolk DHA levels, and deceases n-6/n-3 fatty acid ratio.
INTRODUCTION
Among various marine resources, microalgae has recently received worldwide attention, as a potent source for biodiesel, cosmetics and organic fertilizers. Furthermore, microalgae has also been utilized as a source of protein and fiber for nutritional supplementation in humans, and its application has been extended to domestic livestock (Lum et al., 2013; Gatrell et al., 2014).
Microalgae has an excellent nutritive value. It is a rich source of essential amino acids, vitamins, minerals, carotenoids and fatty acids (Becker, 2004). In addition, microalgae has many potential health promoting effects, such as antioxidant activity, immune stimulating properties, decreasing blood pressure and cholesterol, and promoting growth of beneficial microorganisms in the intestine (Jimenez-Escrig et al., 2001; Hata et al., 2002; Hirahashi et al., 2002; Spolaore et al., 2006; Dvir et al., 2009). The dietary supplementation of microalgae rich in docosahexaenoic acid (DHA) increases DHA content in milk fat of dairy cattle, muscle tissue of pigs, and the egg yolk of layers (Becker, 2004; Sardi et al., 2006; Stamey et al., 2012). Moreover, microalgae in poultry diets can be effectively used as a natural pigment source, both in egg yolk, and in broiler skin (Herber and Van Elswyk, 1996; Sardi et al., 2006).
Microalgae is abundant, has a rapid growth rate and is thought to be an attractive feed additive, because of its digestible protein and long-chain omega-3 fatty acids, which could be substituted for fish oils or flaxseed and canola oils (Trentacoste et al., 2013). Although microalgae has high potential for use as an animal feed additive, researches on the effect of microalgae are insufficient in layers. Therefore, the aim of this study was to investigate the effect of marine microalgae, Schizochytrium with rich DHA on egg production, egg quality, blood profiles, and fatty acid composition of egg yolk in layers.
MATERIALS AND METHODS
Experimental birds and design
All animal-based procedures were in accordance with the Guidelines for the Care and Use of Experimental Animals of Dankook University.
A total of 216 Institut de Sélection Animale (ISA) brown commercial layers (1,835 g±42) were randomly assigned to 3 treatments, with 12 replications of 6 hens in each treatment, during 40 to 46 wks of age. Six layers were housed per 114×50×40 cm wire cage, and were subjected to a photoperiod of 16 h light and 8 h dark/d. Layers were fed mash diets, formulated to supply their nutritional requirements (NRC, 1994) (Table 1). Feed and water were provided ad libitum. Dietary treatments were as follows: i) CON (basal diet); ii) 0.5% microalgae powder (MAP; CON+0.5% Schizochytrium powder), and iii) 1.0% MAP (CON+1.0% Schizochytrium powder). Dried microalgae from Schizochytrium sp. JBF-06 strain was obtained from JINIS Co., Ltd., Wanju, Korea. The fatty acid content of MAP is shown in Table 2.
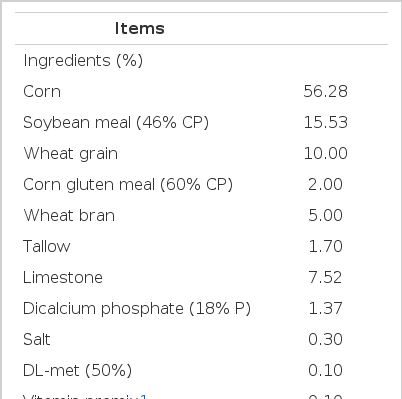
Basal diet composition (as-fed basis)

Fatty acid composition of microalgae (Schizochytrium) powder
Egg production, egg quality and blood profiles
Daily egg production and egg weight were recorded. Forty eight eggs from each treatment (4 eggs/cage) were randomly collected at 40, 43, and 46 wks, and used to determine the egg quality. Haugh unit and yolk color were evaluated, using an egg multi tester (Touhoku Rhythm Co. Ltd., Tokyo, Japan). Eggshell breaking strength was evaluated, using an Eggshell force gauge model II (Robotmation Co., Ltd., Tokyo, Japan); and eggshell thickness was measured, using a dial pipe gauge (Ozaki MFG. Co., Ltd., Tokyo, Japan).
Blood samples were randomly collected from 24 layers in each treatment (2 layers/cage) at 46 wks, using a sterilized syringe and K3EDTA vacuum tubes (Becton Dickinson Vacutainer Systems, Franklin Lakes, NJ, USA). The blood samples were then centrifuged at 3,000 rpm at 4°C for 20 min within one hour of collection, to separate the serum. The concentration of triglyceride and total cholesterol in the serum samples were determined by enzymatic colorimetric methods (Stein et al., 2002), using an automatic biochemical analyzer (RA-1000, Bayer Corp., Tarrytown, NY, USA).
Fatty acid composition of egg yolk
At the end of the experiment (46 wks), 48 eggs were randomly selected from each treatment (4 eggs/cage), and the fatty acid composition of egg yolk was determined according to the modified method of Kim et al. (2003). Briefly, the lipid from the egg yolk was extracted with hexane/isopropanol (3:2 v/v). The extracted lipids were mixed with 0.5 mL of toluene and 2 mL of 5% KOH-MeOH, and heated at 70°C for 8 min; and then 2 mL of 14% BF3-MeOH was added to the above mixture, and heated at 70°C for 2 min. The fatty acid methyl esters (FAME) were extracted with 3 mL of 5% NaCl and 1 mL of hexane. Samples were analyzed for total fatty acids, using an HP 5,890 gas chromatography with a flame ionization detector (Hewlett Packard 5890 Series II, Palo Alto, CA, USA). The FAME were separated using a Supelcowax-10 fused silica capillary column (100 m×0.32 mm×0.25 μm; Supelco, Inc., Bellefonte, PA, USA), with 1.2 mL/min of helium flow. The oven temperature was increased from 220°C to 240°C, at the rate of 2°C/min. Temperatures of the injector and detector were 240°C and 250°C, respectively. The peak of fatty acids were identified by comparing the retention time and peak area of each fatty acid standard, respectively. The content of each fatty acid was expressed as a percentage of the sum of all of the fatty acids analyzed.
Statistical analysis
Data were statistically analyzed by analysis of variance, using general linear model procedure of the SAS program (SAS, 2002), for a completely randomized design. Mean values and standard errors of means are reported. Orthogonal polynomial contrast was conducted to measure the linear and quadratic effects for increasing the MAP levels on all measurements. Statements of statistical significance were based on p<0.05.
RESULTS
Egg production
Layers fed with diets supplemented with MAP during 40 to 43 wks showed no significant difference in egg production, compared with the control treatment. However, layers fed with MAP showed significantly higher egg production during 44 to 46 wks, as dietary MAP increased from 0.5% to 1.0% (linear, p = 0.034); but there was no significant difference in egg weight (Table 3).
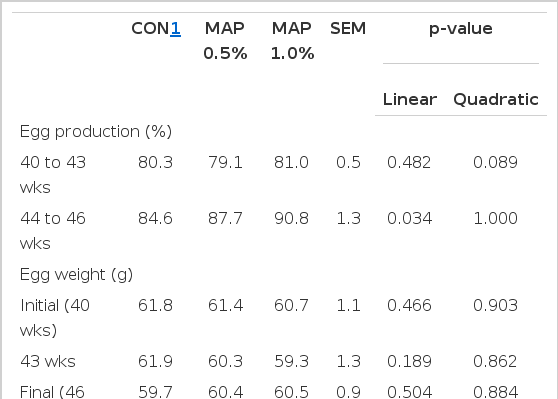
Effects of dietary microalgae (Schizochytrium) supplementation on egg production and egg weight in layers
Blood profiles
Blood triglyceride concentrations increased quadratically in MAP treatment, compared to control treatment (p = 0.034). Similarly, total cholesterol concentrations of blood also increased quadratically (p = 0.039, Table 4).

Effects of dietary microalgae (Schizochytrium) supplementation on blood lipid metabolites in layers
Egg quality
There were no significant differences in Haugh unit and egg breaking strength of layers fed with different levels of MAP in diets. However, the yolk color index was found to quadratically increase, compared with the control group, at 46th wk (p = 0.044). Also, there was a linear effect on eggshell thickness, as dietary levels of MAP increased at 46th wk (p = 0.003, Table 5).

Effects of dietary microalgae (Schizochytrium) supplementation on egg quality
Egg yolk fatty acid
The fatty acid composition of egg yolk is shown in Table 6. Palmitic acid (C16:0), DHA (C22:6n-3) and saturated fatty acid (SFA) compositions increased linearly, in layers fed with MAP (p = 0.044, p = 0.010, and p = 0.011, respectively). The eicosapentaenoic acid (EPA; C20:5n-3) composition was also found to be higher in 0.5% MAP treatment, than in CON and 1.0% MAP treatments; and it had quadratic (p = 0.008) effect. Linoleic acid (C18:2n-6), n-6 fatty acid, unsaturated fatty acid (USFA) composition and USFA/SFA ratio decreased in layers fed with MPA (linear, p = 0.013, p = 0.013, p = 0.002, and p = 0.005). In addition, n-6/n-3 fatty acid ratio also decreased as dietary levels of MAP increased, and it had a linear (p = 0.023) effect.
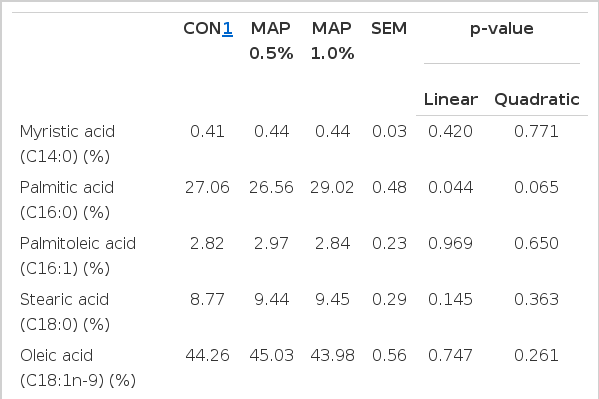
Effects of dietary microalgae (Schizochytrium) supplementation on fatty acid composition of egg yolk
DISCUSSION
Some species of the genus Thraustochytrium, such as Schizochytrium, are known to contain large amounts of DHA in their cytoplasm, and have been used as alternative sources for commercial DHA production. In the present study, we confirmed that Schizochytrium contains a high DHA concentration of about 37.6% of total fatty acids (Table 2). We hypothesized that the supplementation of MAP containing high DHA level to layer diets would result in positive effects on blood lipid metabolites and egg yolk fatty acid composition.
The addition of 0.5% and 1.0% MAP resulted in dose-dependent changes of egg production (44 to 46 wks); but did not significantly affect egg weight. It is well documented that dietary sources of n-3 polyunsaturated fatty acids, such as fish oil, linseed oil and canola oil, have no effect on egg production in layers (Hargis et al., 1991; Mazalli et al., 2004). The results of research on layer’s performance responses to dietary microalgae supplementation is inconsistent. Zheng et al. (2012) reported that inclusion of microalgae (Chlorell vulgaris) in the laying hen feed had positive effects on egg production. However, Lemahieu et al. (2013) found that egg production was not significantly affected by supplementation of omega-3 rich microalgae (Phaeodactylum tricornutum, Nannochloropsis oculata, Isochrysis galbana, or Chlorella fusca) to the layers. Since there is currently a lack of useful information regarding the use of microalgae Schizochytrium in layers, our study may not be able to completely explain its improvement of egg production. However, microalgae are generally known to contain many essential amino acids, carotenoids, vitamins, minerals, and USFA; and have received much attention as a potential natural antioxidant. In addition, feeding microalgae (Chlorella vulgaris) to layers also results in increased beneficial microbial diversity in the crop and ceca (Janczyk et al., 2009). Therefore, these nutritional factors and change in the intestinal microbial community seem to be involved in some of the metabolism related to egg production. The findings of our current study imply that MAP from Schizochytrium in the diet may have an improving effect on layer performance; though no significant differences were seen in egg weights between the MAP groups and control. In the present study, reduced serum levels of triglycerides and cholesterol were observed in layers fed with MAP in the diet. The underlying mechanism in relation to lipid metabolism was unclear, and the active ingredients responsible for the activity have not been identified. However, DHA from fish oil and microalgal sources has been found to have beneficial effect on risk factors for cardiovascular disease, including reductions in blood triglyceride levels and in the triglyceride: HDL-cholesterol ratio, as well as shifts towards moderate elevations in HDL-cholesterol levels (Holub, 2009). In a recent study, Chen et al. (2011) suggested the possibility that DHA from a microalgal source can inhibit 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase activity, which reduces cholesterol synthesis, and therefore lowers the plasma cholesterol level. The HMG-CoA reductase is a rate-limiting enzyme in cholesterol biosynthesis; and its inhibition can lead to reduction in plasma cholesterol. In addition, numerous studies indicate that dietary fish oil lowers the level of total cholesterol, low-density lipoprotein and triglycerides. Also, the other probable cause is that decreased levels of blood triglyceride and cholesterol found in MAP groups may be associated with polysaccharides in microalgae. Marine algae contains large quantities of soluble and insoluble dietary fibers (Lahaye and Jegou, 1993), which have been reported as successfully reducing blood lipids (Ara et al., 2002; Werman et al., 2003). Therefore, decreased blood triglyceride and total cholesterol concentration in this study were due to the inclusion of dietary MAP, related to the DHA or polysaccharides content in MAP.
Herber and Van Elswyk (1998) indicated that eggs from hens fed with n-3 fatty acid enriched marine microalgae had significantly increased yolk redness values, as compared with controls. Enhancement of yolk redness values most likely reflects the deposition of marine microalgae carotenoids. Some microalgae ingredients used for egg enrichment with polyunsaturated n-3 fatty acids are natural sources of carotenoids, and their use in layer diets increases the yolk color index. Actually, Schizochytrium algae used in this study had a yellow-dark orange color, although we did not measure the pigments in MAP. Aki et al. (2003) reported high levels of β-carotene and xanthophylls, including canthaxanthin and astaxanthin, in Schizochytrium; and Kotrbacek et al. (2013) confirmed that carotenoids are efficiently deposited in egg yolk, when added to poultry diets. Therefore, addition of MAP into layer diet efficiently increased yolk color, which gives an opportunity for egg yolk pigmentation. Our study also showed that supplementation of MAP had a beneficial effect on the eggshell thickness. However, until now there was no scientific evidence to support the fact that microalgae can affect the eggshell thickness. In general, marine algae are rich natural sources of trace minerals. We presume that our findings were related to organic minerals contained in MAP influencing the eggshell thickness. The use of microalgae in mineral metabolism in layer needs further investigation
In the present study, n-3 fatty acid concentrations in eggs were found to be significantly increased by MAP supplementation. Considering that MAP contains high DHA, and dietary fat is directly reflected in the fatty acid profiles of egg, the rise in n-3 fatty acid levels is easily understood. Oleaginous marine microorganism such as Schizochytrium has recently become available as a source of fish oils, for various functional food fortifications. Therefore, some of the strains, including Schizochytrium, have been mainly used as poultry feed additives to produce omega 3-enriched eggs (Chin et al., 2006; Lemahieu et al., 2013). Similar to previous reports, MAP leads to increased total DHA in the eggs. Polyunsaturated fatty acids intake including n-3 and n-6 fatty acids, has beneficial effects on both animal and consumer health. However, some limited studies imply that excessive intake of n-6 relative to n-3 fatty acids was associated with an increased in cardiovascular heart disease or inflammatory diseases (Simopoulos, 2003; Hibbeln et al., 2006). Since consuming a higher amount of n-6 fatty acids has been closely associated with negative health impact, attempts have been made to manipulate the fatty acid profile in human diet, using several fat sources (Huang et al., 1990; Shin et al., 2011). Dietary n-6 fatty acid ratio, relative to n-3 fatty acid, was recommended to be decreased, for ideal nutritive value in humans (Gerster, 1998); but it was lower in our study, in both the 0.5% MAP and 1.0% MAP groups (8.20 and 6.08, respectively). On the other hand, control group showeda higher value (11.36) than these groups. It can thus be proposed that MAP fed to layers is a feeding practice that is able to decrease the n-6 to n-3 fatty acid ratio in their eggs. Total SFA significantly increased in a linear manner in egg yolks of layers fed with MAP diets. An increase in proportion of SFA seems mainly to be associated with higher n-6 fatty acids and lower C16:0.
In conclusion, supplementation of MAP in the diet of layers had a beneficial effect on egg production, egg weight, yolk color and blood lipid profiles of layers, in comparison with the control group. In addition, MAP could enhance the DHA and EPA concentrations in eggs, and decrease n-6 fatty acid or n-6/n-3 fatty acid ratio.
ACKNOWLEDGMENTS
The present research was conducted by the research fund of Dankook University in 2014.