Effects of Cellulase Supplementation on Nutrient Digestibility, Energy Utilization and Methane Emission by Boer Crossbred Goats
Article information
Abstract
This study examined the effect of supplementing exogenous cellulase on nutrient and energy utilization. Twelve desexed Boer crossbred goats were used in a replicated 3×3 Latin square design with 23-d periods. Dietary treatments were basal diet (control, no cellulase), basal diet plus 2 g unitary cellulase/kg of total mixed ration dry matter (DM), and basal diet plus 2 g compound cellulase/kg of total mixed ration DM. Three stages of feeding trials were used corresponding to the three treatments, each comprised 23 d, with the first 14 d as the preliminary period and the following 9 d as formal trial period for metabolism trial. Total collection of feces and urine were conducted from the 4th d of the formal trial, and gas exchange measures were determined in indirect respiratory chambers in the last 3 d of the formal trial. Results showed that cellulase addition had no effect (p>0.05) on nutrient digestibility. Dietary supplementation of cellulase did not affect (p>0.05) N intake and retention in goats. Gross energy (GE) intake, fecal energy and urinary energy excretion, heat production were not affected (p>0.05) by the cellulase supplementation. Total methane emission (g/d), CH4 emission as a proportion of live weight or feed intake (DM, organic matter [OM], digestible DM or digestible OM), or CH4 energy output (CH4-E) as a proportion of energy intake (GE, digestible energy, or metabolizable energy), were similar (p>0.05) among treatments. There was a significant (p<0.001) relationship between CH4 and live weight (y = 0.645x+0.2, R2 = 0.54), CH4 and DM intake (y = 16.7x+1.4, R2 = 0.51), CH4 and OM intake (y = 18.8x+1.3, R2 = 0.51) and CH4-E and GE intake. Results from this study revealed that dietary supplementation of cellulase may have no effect on nutrient digestibility, nitrogen retention, energy metabolism, and methane emission in goat.
INTRODUCTION
Cellulase is believed to be effective in improving nutrient digestibility of ruminants, especially rumen degradability of fiber. Ballard et al. (2003) reported that dietary addition of compound cellulase (CC) increased the apparent digestibility of dry matter (DM), organic matter (OM), and neutral detergent fiber (NDF) of dairy cows. Bilik and Łopuszańska-Rusek (2010) reported that the addition of exogenous cellulase may increase total volatile fatty acid (VFA) of rumen fluid of dairy cows without disturbing the VFA profile. Rumen VFA is the energy source for ruminants, and therefore it could be deduced that exogenous cellulase might affect energy metabolism of ruminants. There have been considerable research on the energy metabolism of different species of ruminants, but no research on the effect of exogenous cellulase supplementation on energy metabolism of ruminants has been reported. As the evidence for nutrient digestibility being elevated by exogenous cellulase accumulated (McAllister et al., 2000; Tang et al., 2013; Vijay Bhasker et al., 2013), this study hypothesized that energy utilization could also be affected by the dietary addition of exogenous cellulase. Therefore, the objective of this study was to determine the effects of exogenous cellulase on nutrient utilization by Boer crossbred goats.
MATERIALS AND METHODS
All animals in this study were cared for according to the standards set by the Chinese guidelines for animal welfare and the experimental protocol was approved by the Animal Care and Use Committee of the Chinese Academy of Sciences.
Animals and diets
Twelve castrated Boer crossbred goats (Boer×Jianchang Black), average body weight (BW) 20±1.51 kg and aged around 10 months, were used in a four replicate 3×3 Latin square design. These goats were purchased from farmers around Chengdu. The sexual maturity of this species is at 7 to 8 months of age, and the growing age until 12 months of age. They were housed individually in metabolism cages designed to separately collect feces and urine. Before the experiment, goats were dewormed for internal parasites with albendazole (Yakang Animal pharmacy, Weifang, Shandong, China) at 12 mg/kg BW. There were three experimental periods corresponding to three treatments. Each stage comprised 21-d periods, with the first 14 d as the adaptation period and the following 9 d as the measurement period including 6 d in crates and 3 in chambers.
From d 1 to d 6 of the formal trial, goats were kept in metabolism cages for total collection of feces and urine. On d 7 of the formal trial, the goats were transferred into respiratory chambers for adaptation; on d 8 and d 9, the goats were subject to gas exchange measurements in respiratory chambers for measuring oxygen consumption, carbon dioxide output, and methane output. Total feces and urine were collected daily both in chambers and in metabolism cages.
The experimental diet comprised of 30% concentrate, 35% alfalfa and 35% Chinese wildrye (Table 1), and was formulated to provide adequate digestible energy (DE) and crude protein (CP) for a 20-kg goat gaining 100 g/d of body weight according to NRC (1981). Dietary treatments were basal diet (control, no cellulase), basal diet plus 2 g unitary cellulase (UC)/kg of ration DM, and basal diet plus 2 g CC/kg of ration DM. The enzymes (UC and CC) used in this study were commercial preparations of fungal extracts (Youter Biotechology Shanghai Co., Ltd., Shanghai, China). The UC additive contained 10,000 IU/g of cellulase (endoglucanase, EC 3.2.1.4) activity, while CC additive contained 7,000 IU/g of cellulase (endoglucanase, EC 3.2.1.4) and 5,000 IU/g of xylanase (1, 4-β-xylanase, EC 3.2.1.8) activity. Cellulase activity was determined at 39°C and pH 6.0 using carboxymethyl cellulose (catalog no. C-5678; Sigma Chemical Co., St. Louis, MO, USA) as substrates and xylanase activity was determined at 50°C and pH 5.5 using birchwood xylan (catalog no. X-0502; Sigma Chemical Co., USA) as substrates. The enzyme additive (i.e., 2 g of the enzyme additive per kg of ration DM for UC and CC treatment) was first diluted in 40 mL of water, and the dilution was added to the concentrate at the time of mixing, usually within 1 h before feeding. An equal amount of water (40 mL/kg of concentrate DM) was added to the control diet. Experimental diets were fed to goats for ad libitum intake twice daily with equal amounts at 1,000 and 1,600 h, and fresh water was available freely at all times.
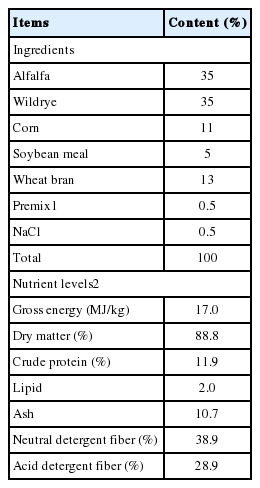
The ingredient and chemical composition of the diet (DM basis)
Measurements
Live weight was recorded for 3 consecutive days in the morning before feeding at the beginning and end of each period. Feed intake for individual goats was recorded daily during the 9-d formal trial in each period. Experimental diets, dietary ingredients, and refusals for individual goats were sampled at the beginning and end of formal trial of each period and composited by period. Periodical refusals, experimental diets, and ingredients were stored at −20°C until analyzed. Feces were collected daily in wire-screen baskets placed under the floor of metabolism crates, and urine was collected through a funnel into plastic buckets. Urine and feces were weighed daily. Urine was acidified with 10% HCl (vol/vol, 5% of urine), and feces were preserved with 10% formaldehyde (vol/vol, 3% of feces) and thoroughly mixed. Urine and feces were sub-sampled daily after acidification and preservation, and stored frozen at −20°C for later analysis.
Feed offered, refusals, and fecal samples were dried in a forced-air oven at 65°C for 48 h, then ground to pass through a 1-mm screen, and were analyzed for DM, gross energy (GE) (AOAC, 1990), and NDF (filter bag technique; China Agricultural University). Urine samples were assayed for N and GE concentrations.
For calorimetry measures, goats were moved into indirect open-circuit calorimetry chambers. The concentration of O2, CO2 and CH4 were analyzed using gas chromatography. Before the gas exchange measurements, validity and accuracy of expired CO2 and inspired O2 flows were checked with ethane combustion with the same flow rates as used during measurements. Before each test, analyzers were calibrated with reference gases: 99.99% O2, 99.99% CO2, and 99.99% CH4. Temperature and humidity in the chambers was maintained at 23°C and 50% to 55%, respectively.
Calculations
Metabolizable energy intake (MEI) was calculated by the difference between GE intake (GEI) and energy losses in feces (FE), urine (UE), and CH4 (CH4E)
Heat production (HP) was estimated based on the Brouwer (1965) equation:
CH4 contains 39.5388 kJ/L energy as reported by Brouwer (1965). Retained energy (RE) was determined as the difference between MEI and HP.
Statistical analyses
Data were analyzed using the MIXED procedure (SAS Institute Inc., Cary, NC, USA; Littell et al., 1996). Goat was the experimental unit for all variables. The full general linear model (GLM) model included the fixed effects of square, period nested within square, treatment (control, UC and CC enzyme), sampling time (day), and the interaction of treatment and sampling time. For all data, if the interaction of treatment by sampling time was not significant (p>0.05), the fixed effect of sampling time and its related interaction were removed from the full GLM model. Sampling time was a repeated effect in the model. Goat nested within square was used in the random statement. Differences were declared significant at p≤0.05, and a tendency to significance was declared at 0.05<p≤0.10. Means were separated using Duncan’s multiple contrasts.
RESULTS AND DISCUSSION
Nutrient digestibility
There was no significant difference (p>0.05) in live weight, live weight gain and DM intake among treatments. Digestibility of DM, OM, CP, crude fat, NDF and acid detergent fiber (ADF) were not affected (p>0.05) by cellulase treatments (Table 2).
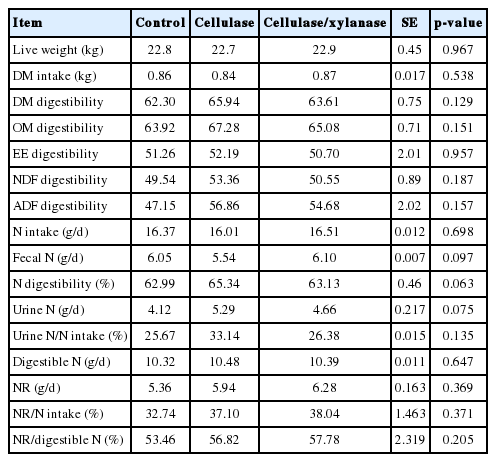
The effects of enzyme treatments on apparent nutrient digestibility and N metabolism in growing goats
There are controversial reports on total tract digestibility of DM and OM or both, following cellulase treatments. Some researchers observed a positive effect of treatment with exogenous cellulase on DM or organic digestibility (Lewis et al., 1996; Rode et al., 1999) while some other studies observed no effect (Lewis et al., 1999; McAllister et al., 2000; Yang et al., 2000). However, present results found that cellulase addition had no effect on DM and OM digestibility. Some researchers found that NDF and ADF digestibility increased following cellulase treatments (Lewis et al., 1999; McAllister et al., 2000), but these results were not observed in this study. The contradictory reports on the effect of cellulase on nutrient digestibility may be due to the digestion characteristics of the rations. Yang et al. (2000) noted that exogenous cellulase improved digestibility of low quality rations, but no effect was evident with high quality rations. This viewpoint still needs confirmation, because O’Connor et al. (2007) found that nutrient digestibility even decreased when cellulase was added to high quality forage diets of horses. Therefore, the effect and mechanisms of exogenous cellulase on nutrient digestion are still incompletely understood. Yang et al. (1999) speculated that improved digestibility caused by exogenous cellulase was due to improved microbial colonization. Wang et al. (2001) and Morgavi et al. (2004) found associative effects between exogenous cellulase and cellulase from rumen micro-organisms. Therefore, exogenous cellulase may improve nutrient digestibility by improving colonization and by stimulating the endogenous enzyme activity within the rumen. But such improved colonization and stimulated endogenous enzyme activity may be related to forage type or ration type.
Nitrogen balance
There was no difference (p>0.05) on N intake, fecal N and urinary N among the treatments. N retention (NR) and N retention rate (NR/NI) were not affected (p>0.05) by the addition of cellulase (Table 2).
Awawdeh and Obeidat (2011) found no effect on NR of lamb after the addition of cellulase at 20 g/d. Although it has been documented that ruminal microbial protein synthesis could be stimulated by the addition of cellulase (Hristov et al., 1998; Rode et al., 1999), however, this result may not be sufficient to explain the subsequent effect on NR. Gado et al. (2009) added cellulase at the rate of 40 g/d in cow diets (30% concentrate), and found no effect on NR and utilization, also microbial protein synthesis was increased. In the present study, the supplementation of cellulase had no effect on NR in goats which is in agreement with the report of Gado et al. (2009) with cows, but in conflict with McAllister et al. (2000) in lambs.
Effect of exogenous cellulase on energy metabolism
No significant (p>0.05) difference was evident on GE intake, fecal energy and urinary energy excretion, CH4 energy, heat production and RE among treatments (Table 4). The addition of UC and CC could significantly increase the DE/GEI and ME/GEI, but did not affect HP/GEI (Table 4).
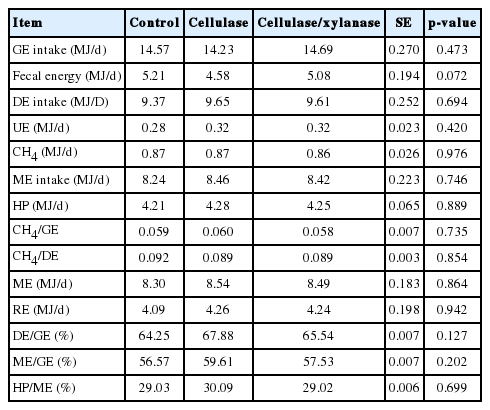
Effects of dietary addition of exogenous cellulase on energy metabolism of growing goats
Few studies concerning the effect of cellulase on energy utilization of livestock have been reported. O’Connor et al. (2007) reported that cellulase supplementation did not improve GE digestibility of horses. In fact, horses consuming the control ration digested a greater (p<0.05) GE than those consuming the ration with cellulase supplementation. Results from the present study showed that GE intake, fecal energy and urinary energy excretion, methane energy and heat production were not affected by the addition of cellulase.
Effect on methane emission
There were no significant differences among dietary treatments on total CH4 emissions (g/d), CH4 emission as a proportion of live weight or feed intake (DM, OM, digestible DM or digestible OM), or CH4 energy output (CH4-E) as a proportion of energy intake (GE, DE, or ME) (Table 3). There was a significant (p<0.001) relationship between CH4 and live weight (y = 0.645x+0.2, R2 = 0.54), CH4 and DM intake (y = 16.7x+1.4, R2 = 0.51), CH4 and OM intake (y = 18.8x+1.3, R2 = 0.51) and CH4-E and GE intake (Table 5 and Figure 1).
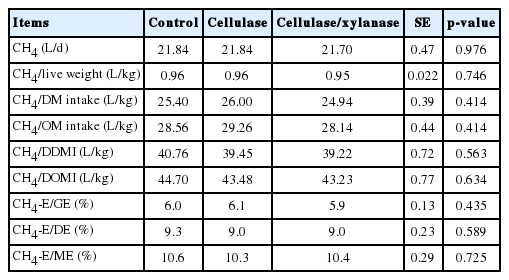
Effects of dietary addition of exogenous cellulase on enteric methane emission in growing goats

The relationships between methane emission and live weight, feed intake or energy intake in growing goats

The linear relationships between methane emission and live weight (A), and between methane energy put and gross energy (GE) intake (B) in growing goats.
Methane emission from ruminants is receiving an increased level of public attention. Different varieties of additives were tested for the mitigation of methane emission. Except for plant extracts, ionophores such as monensin or lasalocid (Guan et al., 2006), supplemental fat (Jordan et al., 2006), halogenated analogues such as bromochloromethane (Abecia et al., 2012) were studied for the possible usage in the mitigation of methane emission from ruminants. But till now, very few studies concerning the effect of cellulase on methane emission have been reported (Tang et al., 2013). Eun and Beauchemin (2007) reported that properly formulated cellulase can lower the acetate-to-propionate ratio, which is thought to be the primary mechanism of methane abatement. Apparently this hypothesis was not supported by our study. Chung et al. (2012) even found negative results, e.g. enteric methane of dairy cow increased linearly with an increasing level of cellulase supplement.
The CH4-E/GE intake ranged from 0.059 to 0.061, which is within recommendations of IPCC (2006) for lambs (0.045) and mature sheep (0.065) for development of Tier 2 emission inventories. There is no recommendation of CH4-E/GE intake for goats in IPCC (2006).
CONCLUSION
On the basis of above results, it may be concluded that cellulase addition had no effect on dry matter intake, nutrients digestibility and N utilization in goats. GE intake, fecal energy and urinary energy excretion, methane emission, heat production were also not affected by the cellulase supplementation.
IMPLICATIONS
The results from this study implicated that exogenous cellulase has no effect on nutrients digestion and N utilization in goats fed on high roughage diet irrespective of sources used. Supplementation of cellulase may not be an effective additive for methane mitigation.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.