Effects of Ecklonia cava as fucoidan-rich algae on growth performance, nutrient digestibility, intestinal morphology and caecal microflora in weanling pigs
Article information
Abstract
Objective
In the present study, role of increasing levels of Ecklonia cava (seaweed) supplementation in diets was investigated on growth performance, coefficient of total tract apparent digestibility (CTTAD) of nutrients, serum immunoglobulins, cecal microflora and intestinal morphology of weanling pigs.
Methods
A total of 200 weaned pigs (Landrace×Yorkshire×Duroc; initial body weight 7.08±0.15 kg) were randomly allotted to 4 treatments on the basis of body weight. There were 5 replicate pens in each treatment including 10 pigs of each. Treatments were divided by dietary Ecklonia cava supplementation levels (0%, 0.05%, 0.1%, or 0.15%) in growing-finishing diets. There were 2 diet formulation phases throughout the experiment. The pigs were offered the diets ad libitum for the entire period of experiment in meal form.
Results
The pigs fed with increasing dietary concentrations of Ecklonia cava had linear increase (p<0.05) in the overall average daily gain, however, there were no significant differences in gain to feed ratio, CTTAD of dry matter and crude protein at both phase I and phase II. Digestibility of gross energy was linearly improved (p<0.05) in phase II. At day 28, pigs fed Ecklonia cava had greater (linear, p<0.05) Lactobacillus spp., fewer Escherichia coli (E. coli) spp. (linear, p<0.05) and a tendency to have fewer cecal Clostridium spp. (p = 0.077). The total anaerobic bacteria were not affected with supplementation of Ecklonia cava in diets. Polynomial contrasts analysis revealed that villus height of the ileum exhibited a linear increase (p<0.05) in response with the increase in the level of dietary Ecklonia cava. However, villus height of duodenum and jejunum, crypt depth, villus height to crypt depth ratio of different segments of the intestine were not affected.
Conclusion
The results suggest that Ecklonia cava had beneficial effects on the growth performance, cecal microflora, and intestinal morphology of weanling pigs.
INTRODUCTION
During the weaning transition undesirable stress accompanied by various changes in the piglet’s gastrointestinal environment result in reduced voluntary feed intake [1], which subsequently increases susceptibility to intestinal dysfunction and weaning related disorders [2]. The widespread use of in-feed antibiotics, at both therapeutic and sub-therapeutic levels, increases public health scares [3]. The underlying mechanisms for the growth-promoting effect of antibiotics are expected to relate to gastrointestinal tract, particularly in the small intestine, at the bacteriological, physiological and immunological level [2]. Dietary manipulations post-weaning have the ability to improve weaning associated with intestinal dysfunction.
Marine algae is a good source of biomass, of which, a few types have been identified as rich sources of structurally diverse bioactive compounds with great pharmaceutical and biomedical potential. Presently these identified types are mainly being used for consumption by humans. It is estimated that two-thirds of the earth’s biomass is comprised of more than 25,000 species of algae [4]. That statistic shows the potential for algae to be used as animal feed additives due to their vast biodiversity [5] and has recently been discussed as a source of bioactive compounds due to their biological properties [6,7]. Ecklonia cava (E. cava) is among one of the types of seaweed, which is found mainly in South Korea, Japan and China and widely used in human food because of their therapeutic effects. It is a highly resilient brown algae that grows in clear waters [8] and is well known for having fucoidans and polyphenolic compounds (phlorotannins). Fucoidans are branched polysaccharide sulfate esters extracted from the cell wall of marine algae, containing l-fucose as one of the major monosaccharides [9] and they have been shown to have antibacterial [6], immunomodulating [10] and antioxidant [11] activities. Previous researches suggested that E. cava can be used as environment friendly substitutes for antibiotics in diets to improve growth in fishes [12] and the immune systems of animals [13]. However, as per our knowledge, not much work has been done to utilize E. cava as feed additive for pigs. In this study, non-antibiotic feed additives are being investigated.
It is hypothesized that the inclusion of E. cava will improve the performance of weanling pigs by modifying the gastrointestinal tract microflora, digestibility and immunity. Therefore, the aim of the present experiment is to evaluate the effect of varying dietary inclusion levels of E. cava on the growth performance, coefficient of total tract apparent digestibility (CTTAD) of nutrients, serum immunoglobulins, cecal microflora and intestinal morphology of weanling pigs.
MATERIALS AND METHODS
The protocol for the present experiment was approved by the Institute of Animal Care and Use Committee of Kangwon National University, Chuncheon, Republic of Korea. The experiment was conducted at the facility of Kangwon National University farm. All the pigs (Landrace×Yorkshire×Duroc) were housed in partially slatted concrete floor pens (pen size 1.90 m×2.54 m) which were equipped with self-feeder and nipple drinker to allow ad libitum access to feed and water.
Ecklonia cava collection and processing
The seaweed E. cava was directly collected from the sea near Jeju Island, washed three times with water to remove the impurities and then dried. The stem was then removed and only leaves were ground to make a particle size of about 100 μm. This 100 μm powder form of E. cava was directly used in the feed. The rate of fucoidan, proportion of monosaccharaides and chemical composition (Table 2) were measured using the previously reported methods [14].
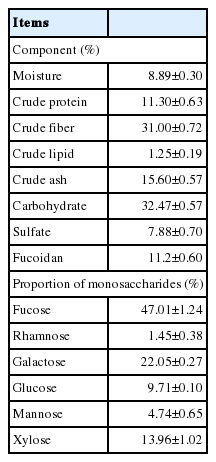
Chemical composition of Ecklonia cava
Animals and experimental design
A total of 200 weaned pigs (Landrace×Yorkshire×Duroc; average initial body weight (BW): 7.08±0.15 kg) were randomly allotted to four treatments on the basis of BW and sex (all male or all female). There were five replicate pens in each treatment with 10 pigs per pen. Treatments were divided by dietary E. cava supplementation levels (0%, 0.05%, 0.1%, or 0.15%) in growing-finishing diets. There were 2 diet formulation phases throughout the experiment (Table 1). The diets were prepared in meal form and were formulated to contain 14.22 MJ/kg metabolism energy (ME) and 1.53% lysine (phase I; Table 1) and 14.02 MJ/kg ME and 14.0 g/kg lysine (phase II; Table 1). All diets met or exceeded the nutrient requirements as recommended by [15].
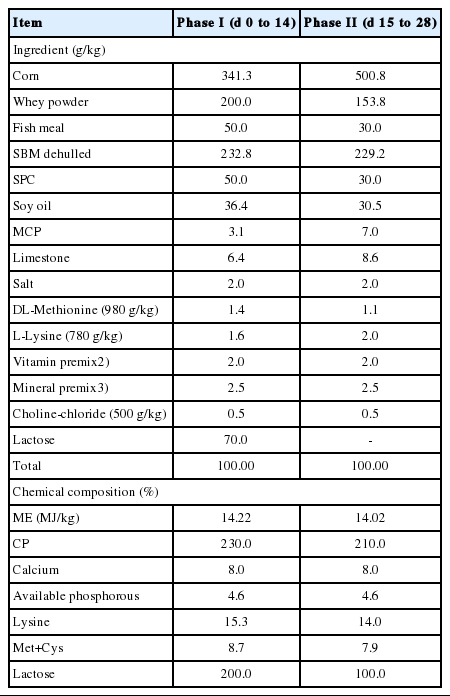
Sample preparation and measurements
The pigs were weighed individually at the beginning of the trial, and then at d 14 and d 28 respectively. Consumption of the feed was recorded at the end of each phase. On the bases of these records, average daily gain (ADG), average daily feed intake (ADFI) and gain to feed ratio (G:F) were calculated.
To evaluate the digestibility, 2.5 g/kg of chromium was added in the diets as indigestible marker and the diets were fed to pigs during last seven days of each phase. Fecal grab samples were collected randomly from four pigs of each pen during the last three days of each phase. Feces were pooled and dried in an air forced drying oven at 60°C for 72 h and ground in a Wiley laboratory mill (Thomas Model 4 Wiley Mill, Thomas scientific, Swedesboro, NJ, USA) using a 1-mm screen for chemical analysis.
To analyze the concentrations of immunoglobulins (IgG, IgA, and IgM), 10 mL blood sample was collected from two randomly selected pigs from each pen at d 14 and d 28 of the experiment. The sampling was done by jugular vein puncture using a disposable vacutainer tube without anticoagulants (Becton Dickinson, Franklin, NJ, USA). The blood samples were then centrifuged (3,000×g for 15 min 4°C) for serum separation which were then stored at −20°C until analysis.
To study the effect of diets on the small intestinal morphology and cecal microflora, pigs from each treatment (two per replicate) reflecting average BW were selected and sacrificed by electrocution at the end of the experiment (d 28). The cecum contents were collected in sterilized plastic bottles and stored at 7°C for bacterial analysis. The intestinal samples from the region of duodenum, jejunum and ileum were also collected after removing their contents and flushing with physiological saline. The samples were submerged in a fixative solution (0.1 M collidine buffer, pH 7.3) containing 30 g/L glutaraldehyde, 20 g/L paraformaldehyde and 15 g/L acrolein and then brought to the laboratory for studying the morphological changes.
Chemical and microbial analyses
Experimental diets and excreta samples were analyzed in triplicate for dry matter (DM, method 930.15; 16), crude protein (CP, method 990.03; 16), ash (method 942.05; 16), calcium and phosphorus (method 985.01; 16). The gross energy (GE) of diets and feces were measured by a bomb calorimeter (Model 1261, Parr Instrument Co., Moline, IL, USA), and chromium concentration was determined with an automated spectrophotometer (Jasco V-650, Jasco Corp., Tokyo, Japan) according to the procedure of [17]. Amino acid composition of feed samples was determined by HPLC (Waters 486, Waters Corp., Milford, MA, USA) after acid hydrolysis [18]. The methionine and cystine were determined following oxidation with performic acid [19]. The concentrations of serum IgG, IgA, and IgM were analyzed using radial immune-diffusion kits (Tripple J Farms, Bellingham, WA, USA).
The microbiological assay of cecal samples was carried out by the procedure suggested by [20]. One gram of mixed content was diluted with 9 mL of Butterfields phosphate buffer solution, followed by further serial dilutions in Butterfields phosphate buffer dilution solution. Duplicate plates were then inoculated with 0.1 mL sample and incubated. The microbial groups enumerated were total anaerobic bacteria (TAB, plate count agar, Difco Laboratories, Detroit, MI, USA), Lactobacillus spp. (MRS agar+0.200 g/L NaN3+0.500 g/L L-cystine hydrochloride monohydrate) and Clostridium spp. (Tryptose sulphite cycloserine agar, Oxoid, Hampshire, UK). The anaerobic conditions during the assay of total anaerobic bacteria and Clostridium spp. were created by using gas-pak anaerobic system (BBL, No. 260678, Difco, Detroit, MI, USA). The microbial populations were log transformed before statistical analysis.
Small intestinal morphology
Three cross-sections for each intestinal sample were prepared after staining with azure A and eosin using standard paraffin embedding procedures. A total of 10 intact and well-oriented crypt-villus units were selected in triplicate for each intestinal cross-section. Villus height was measured from the tip to the villus crypt junction. Crypt depth was measured as the depth between adjacent villi. All morphological measurements (villus height or crypt depth) were made in 10 μm increments by using an image processing and analysis system (Optimus software version 6.5, Media Cybergenetics, North Reading, MA, USA).
Statistical analyses
The data were analyzed as a randomized complete block design using the general linear model procedure of SAS [21]. The linear and quadratic contrasts were used to compare effect of increasing dietary E. cava levels (0, 0.5, 1.0, and 1.5 g/kg). The pen was used as the experimental unit for the analysis of all the parameters. Probability values of ≤0.05 were considered as significant.
RESULTS
Growth performance
As shown in Table 3, during the phase I and II, there was no significant effects of dietary E. cava on ADG, ADFI, and G:F. A linear response to increasing dietary E. cava was demonstrated for the overall ADG (p<0.05). However, dietary E. cava did not affect the overall ADFI and G:F.

Effects of supplemental Ecklonia cava on growth performance of weanling pigs
Digestibility
Polynomial contrasts analysis showed no significant differences in the CTTAD of GE and CP at both the phases (d 14 and d 28), but the digestibility of GE in the second phase linearly improved (p<0.05; Table 4).
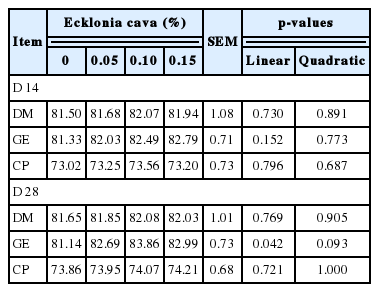
Effects of supplemental Ecklonia cava on apparent total tract digestibility (%) of nutrients in weanling pigs
Cecal microbial population
The Pigs supplemented with E. cava in diets showed no effect on the total anaerobic bacteria populations. However, the population of Lactobacillus spp. was linearly increased (p<0.05; Table 5) in pigs supplemented with increasing E. cava levels in diets. The population of E. coli were linearly reduced (p<0.05; Table 5) and population of Clostridium spp. tended to decrease (p = 0.077) linearly in E. cava supplemented pigs.
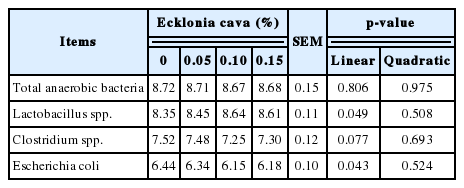
Effects of supplemental Ecklonia cava on caecal microbial populations (Log10 CFU/g) in weanling pigs (d 28)
Serum immunoglobulins
Increasing E. cava level in the diets had no significant effect on serum level of IgG, IgA, and IgM (Table 6) in both phases.

Effects of supplemental Ecklonia cava on serum immunoglobulins (mg/mL) of weanling pigs
Intestinal morphology
The diets supplemented with increasing levels of E. cava had higher villus height (p<0.05; Table 7) in ileum, but villus height was not affected in other sections of intestine. Crypt depth and villus height to crypt depth ratio was unaffected (Table 7) by supplementation of E. cava in diets.
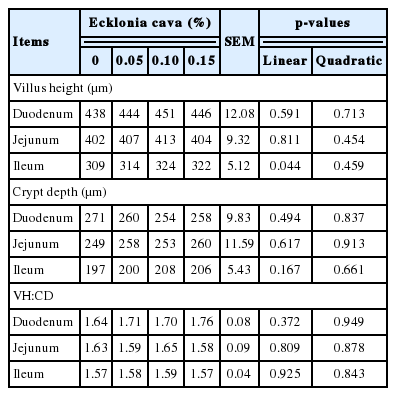
Effects of supplemental Ecklonia cava on small intestinal morphology of weanling pigs (d 28)
DISCUSSION
In the recent years, efforts are being made to select new alternatives for feed additives. Seaweeds due to their enormous biodiversity and availability, have the potential to be used for this purpose [4,5]. A few of the seaweeds have been tried as feed additives to some extent [22,23], however, the focus of the present study to evaluate the effect of a rich fucoidan (E. cava) supplementation in weanling pigs was significant and meaningful. There was a linear increase in the ADG of the weanling pigs with increasing levels of E. cava. This is in line with the earlier reports of [24] who reported inconsistent, but minimal, performance improvement and increase in ADFI after feeding different seaweeds in pig diet. In contrast, [25] reported the addition of rich fucoidan seaweed to well digestible diets did not enhance performances of weaned piglets. This variation in the results may be due to difference in the seaweed variety and the concentration that was used in the diets. In the present study, ADG was observed to be almost 6% higher in 1 g/kg E. cava supplemented diets in comparison to the control diet. The earlier study of [26] reported a 10% increase in the BW gain when the diet was supplemented with algae. The greater ADG in pigs fed the E. cava in diets in the present study might be associated with the improvement in GE digestibility and villi height.
In the present study, the digestibility of GE in the second phase was improved. This is in line with the earlier reports of [27] that reported higher digestibility of GE in rich fucoidan seaweed supplemented diets. Increase in the digestibility in the present study, was also accompanied by a slight increase in villi height. A critical role of villi height in the absorption of intestinal nutrients has been reported [28]. Thus, it is more likely that change in the height of villi could be under the influence of the E. cava in the diets that might be responsible for the increase in nutrient uptake and thus resulted in higher digestibility. [29] studied the nutrient digestibility after dietary supplementation of polysaccharides and found a significant interaction between laminaran and fucoidan supplementation on digestibility of GE. Improvement in the digestibility of nutrients and growth performance could be the result of the decrease of harmful bacteria such as E. coli and the increase of friendly bacteria such as Lactobacillus in weanling pigs’ microbiota. Lactobacilli are able to produce polysaccharide depolymerases and glycosidases, which can improve the digestibility of nutrients by degrading the structural polysaccharide in plant cell wall [30].
In the present study, supplementation of increasing levels of E. cava did not affect the immune status of the piglets. It has been previously reported that polysaccharides have immunomodulatory activities, and the presence of polysaccharides such as fucoidan in E. cava would most likely be responsible for the immunomodulatory activities [31]. However, the significant effect on immunoglobulins observed in previous work could not be confirmed in the current experiment, it might be because of low dosage of E. cava and health status of piglets in this study.
Ecklonia cava, a good source of Fucoidan, a sulfated polysaccharides [27] contains almost 10% of dry mass of total algae weight. In the present study increasing level of E. cava supplementation reduced the Clostridium spp. and E. coli population. This could be due to the presence of sulfated polysaccharides that are well known for their bacterial and viral inhibiting activities [22]. In previous studies, [6] also reported a promising reduction in the coliform populations in the cecum and colon of weaning pigs after supplementing fucoidan extract in diets. Moreover, in earlier studies, [32] reported that polysaccharides can act as prebiotics. Phlorotanin is another important ingredient of E. cava with promising effects against E. coli [33]. In this study, the number of Lactobacillus spp. increased linearly. This could be due to the prebiotic effects of E. cava that may have stimulated the growth of beneficial bacteria in the digestive track and would have exerted growth promoting and health improving effects on the weanling pigs. Moreover, the ratio between these bacterial groups has traditionally been considered as an index of gut health, with a high index related to a greater resistance to intestinal disorders [34]. Therefore, E. cava may offer a dietary means to modulate the gut environment to adjust the intestine microbiota and decrease the risk of diarrhoea in the absence of in-feed antibiotics.
In the present study the villus height in Ileum was linearly improved with increasing levels of E. cava supplementation in diets. The linear increase might have aided the nutrient digestion by enhancing the surface area for absorption. The epithelial turnover and villus height have a direct correlation with the nutrient absorption. A similar result was also reported in the previous study when pigs were offered seaweed extracted polysaccharides (laminaran and fucoidan), the villus height and villus height to crypt depth ratio improved [35]. The higher cecal lactobacillus in probiotic-supplemented diet indicated greater villus height and villus height to crypt depth ratio in the different small intestinal segments resulted in a greater absorption surface and greater digestibility of nutrients such as crude protein, dry matter and GE [36]. Moreover, the suppression of pathogenic bacteria by favorable bacteria results in the better villus height and villus height to crypt depth ratio [36]. The aid for the favorable microorganisms and hindered growth of unfavorable microorganisms could be one of the reasons behind the better gut morphology.
CONCLUSION
In conclusion, effects of E. cava on the digestibility of nutrients and intestine morphology were minimal; however, it had a great effect on growth performance and intestinal microbiota. Trends for increased digestibility and villi height will encourage us to try other levels of E. cava as effective feed additive on pigs in the future studies.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.