Genetic effects of sterol regulatory element binding proteins and fatty acid-binding protein4 on the fatty acid composition of Korean cattle (Hanwoo)
Article information
Abstract
Objective
This study identifies single-nucleotide polymorphisms (SNP) or gene combinations that affect the flavor and quality of Korean cattle (Hanwoo) by using the SNP Harvester method.
Methods
Four economic traits (oleic acid [C18:1], saturated fatty acids), monounsaturated fatty acids, and marbling score) were adjusted for environmental factors in order to focus solely on genetic effects. The SNP Harvester method was used to investigate gene combinations (two-way gene interactions) associated with these economic traits. Further, a multifactor dimensionality reduction method was used to identify superior genotypes in gene combinations.
Results
Table 3 to 4 show the analysis results for differences between superior genotypes and others for selected major gene combinations using the multifactor dimensionality reduction method. Environmental factors were adjusted for in order to evaluate only the genetic effect. Table 5 shows the adjustment effect by comparing the accuracy before and after correction in two-way gene interactions.
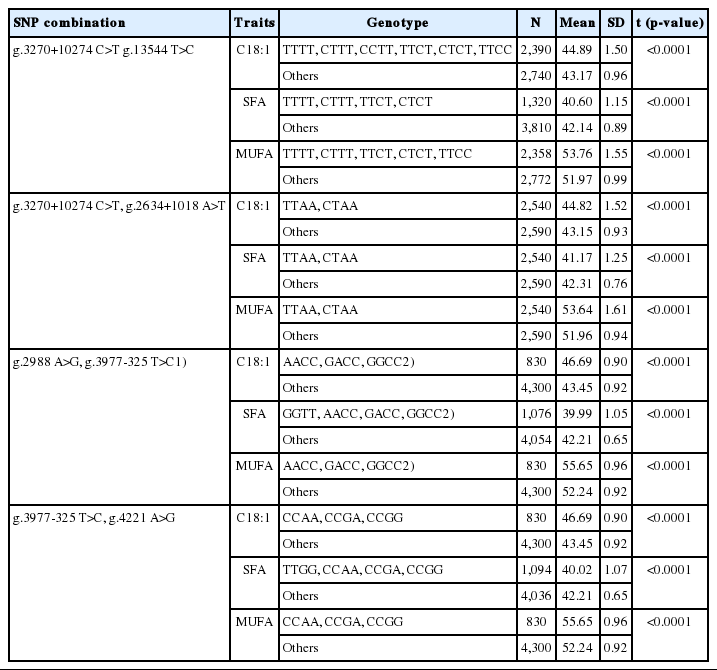
Superior genotypes of four-SNP combinations selected by fatty acids

Superior genotypes of three-SNP combinations selected by MS
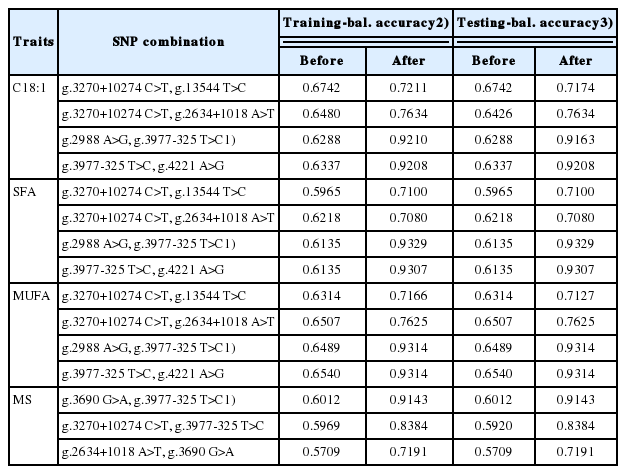
Adjustment effect by comparing accuracy before and after correction in two-SNP combinations
Conclusion
The g.3977-325 T>C and (g.2988 A>G, g.3977-325 T>C) combinations of fatty acid-binding protein4 were the superior gene, and the superior genotype combinations across all economic traits were the CC genotype at g.3977-325 T>C and the AACC, GACC, GGCC genotypes of (g.2988 A>G, g.3977-325 T>C).
INTRODUCTION
In recent years, Korea’s food consumption has increasingly focused on food quality as a result of the public’s favorable perception of well-being. Because meat generally requires high quality, many studies have focused on ways to produce high-quality meat. In general, consumers’ preferences vary according to the taste and meatiness of beef, which are influenced by the fatty acid composition (FAC) and many other factors. Therefore, this study identifies the single-nucleotide polymorphisms (SNP) or gene combinations that influence the flavor and quality of Korean cattle (Hanwoo).
According to the literature, meatiness is determined by marbling score (MS), meat color, and texture, which are closely related to the tenderness, flavor, and juiciness of beef [1–3]. Among these, the flavor of beef is influenced by the content of its unsaturated fatty acids [4–6], and the oleic acid (C18:1) content of Hanwoo is particularly high in unsaturated fatty acids [7–9]. In addition, oleic acid accounts for more than 80% of the monounsaturated fatty acids (MUFAs) and is a key determinant of beef flavor [10,11]. Therefore, four traits (C18:1, saturated fatty acids [SFAs], MUFAs, and MS) have been found to be associated with the flavor and quality of Hanwoo. The routes for synthesizing fatty acids can be divided into two types. Many types of gene contribute to the synthesis of fatty acids. First, chylomicron and very-low-density lipoprotein produced in the small intestine and liver are transformed by the lipoprotein lipase (LPL) gene in the form of fatty acids (Ed- you have not used this fatty acid [FA] acronym in most other places in the paper) [12,13]. Second, Malonyl-CoA, which is a substrate of the biosynthesis of fatty acids converted into 18:0-CoA by the fatty acid synthase (FASN) gene and the stearoyl-CoA desaturase (SCD) gene, converts stearic acid (C18:0) and palmitic acid (C16:0) into oleic acid (C18:1), which is an MUFA of the w-9 line [11,14,15]. In addition, a number of SNP combinations influencing the quality of Hanwoo have been found using the SCD, FASN, and LPL genes [16–18].
This study investigates how major gene combinations and genotypes using fatty acid-binding protein4 (FABP4), which plays a major role in carrying fatty acids and which changes throughout two synthetic pathways in cells [19], and using sterol regulatory element binding proteins (SREBPs), which are transcriptional regulatory factors that influence the biosynthesis of fatty acids, affect the flavor of beef through its FAC and MS [20,21]. In addition, 8 genes (2 of SREBPs and 6 of FABP4) were identified as superior to the remaining 23 genes (10 of SREBPs and 13 of FABP4) based on a test of FAC and economic traits [22]. In particular, as to the SNP selection on FABP4 (Genbank Accession No. NC_007312.4) and SREPBs (Genbank Number NC_007317.4), we have selected the 8 SNPs out of the total of 23 SNPs in our paper based on the result of the study [22] where only 8 SNPs showed polymorphism when 23 SNPs that showed base mutations across all intron and exon on genomic DNA were applied to the Hanwoo.
Environmental factors were adjusted first to examine only the genetic effects and verify the adjustment effects by comparing the accuracy before and after correction. Then, the SNP Harvester method was used to identify major gene combinations developed in Yang et al [23]. However, the SNP Harvester method was applied only to binary data in the case-control study. Therefore, this study’s measures of the four traits were converted into binary values by using k-means cluster analysis. Because genotypes for gene combinations could not be investigated using the SNP Harvester method, a multifactor dimensionality reduction method was employed to identify genotypes in gene combinations [24].
MATERIALS AND METHODS
Animals and phenotypes
Hanwoo cattle were bred in Gyeongbuk, Korea and 513 animals, the progeny of 18 sires, were used in this study. In general, they were weaned and castrated at 6 months of age, fed with growth stage feed for 18 months, and then fed a highly concentrated diet in their last 6 months. All steers were slaughtered at 941±72 days of age, and MS was measured 24 h after slaughter.
Total lipids were extracted from approximately 500 mg of m. longissimus dorsi muscle with chloroform/methanol (2:1, v/v) according to Folch et al [30]. Extracted lipids were then methylated based on O’Keefe et al [26] by using sodium methylate. The extract was then filtered through filter paper in a water bath (40°C). The filtrate was mixed with distilled water, and then a layer of methanol and water was removed. After the removal of chloroform and lipid layers using nitrogen gas, the sample was treated with BF3-methanol (14%) and subjected to trans-methylation at 65°C. Fatty acid content was analyzed using gas-chromatography (PerkinElmer, Inc., Waltham, MA, USA).
Genomic DNA and SNP genotyping
Total genomic DNA was extracted from the longissimus muscle using the LaboPass TM Tissue Mini kit (Cosmo Genetech, Seoul, Korea). Two polymorphic SNPs of SREBPs and six polymorphic SNPs of the FABP4 gene in GenBank were genotyped according to Oh et al [27]. Primers for amplifications and extensions were designed for the single-base extension (Ed- this acronym is not used anywhere in the paper) for genotyping polymorphic sites [33] using forward, reverse, and extension primer sequences [27]. Reactions of the primer extension were performed using the SNaPshotddNTP Primer Extension Kit (Applied Biosystems, Foster City, CA, USA). One unit of shrimp alkaline phosphatase was added to the reaction mixture, which was then incubated for 1 h at 37°C, followed by 15 min at 72°C for enzyme inactivation, to clean the primer extension reaction. DNA samples containing extension products and the Gene-scan 120 LIZ size standard solution were added to Hi-Di formamide (Applied Biosystems, USA) in accordance with the manufacturer’s recommendations. The mixture was incubated for 5 min at 95°C, followed by 5 min on ice, after which electrophoresis was conducted using the ABI PRISM 3130XL GeneticAnalyzer. The analysis was made using GeneMapper v4.0 (Applied Biosystems, USA).
Adjust model and Statistical analysis
Genetic factors influencing the economic traits associated with the flavor and quality of Hanwoo were examined. These economic traits included the breeding farm and age as environmental factors, and SNPs as a genetic factor [29]. To identify the genetic factors influencing economic traits, a model adjusted for environmental factors such as the breeding farm and age was used:
Where yk is a phenotype, μ the overall mean of each trait, agek the age of the kth Hanwoo,
A simple form of the above equation is
Where μ is the total mean vector, Eα the environmental effect (E is an explanatory variable matrix including the environmental factor, and α is a regression coefficient vector representing the effect of environmental factors), and Gβ the genetic effect (G is an explanatory variable matrix including the genetic factor, and β is a regression coefficient vector representing the effect of genetic factors).
The effects α̂ of environmental factors such as age and the breeding farm were examined using the above model, and the economic trait Z was determined for a pure genetic factor to compensate for the effects of environmental factors on estimates. Here the equation was
Where Z is an adjusted value which eliminates the environmental factors. In this study, we use Z as a pure genetic factor for statistical analysis.
SNP harvester
The SNP harvester method can be used to identify major genotype combinations associated with human diseases and economic traits of livestock [23]. This method selects several SNPs linked to economic traits as combinations and then repeats the process to increase scores by changing one of the unchosen SNP-to-SNP combinations. The score function can use the x2 statistic, classification accuracy, and the B-statistic. In this study, the x2 statistic with 3k-1 degrees of freedom was used. The procedure was as follows (Figure 1):
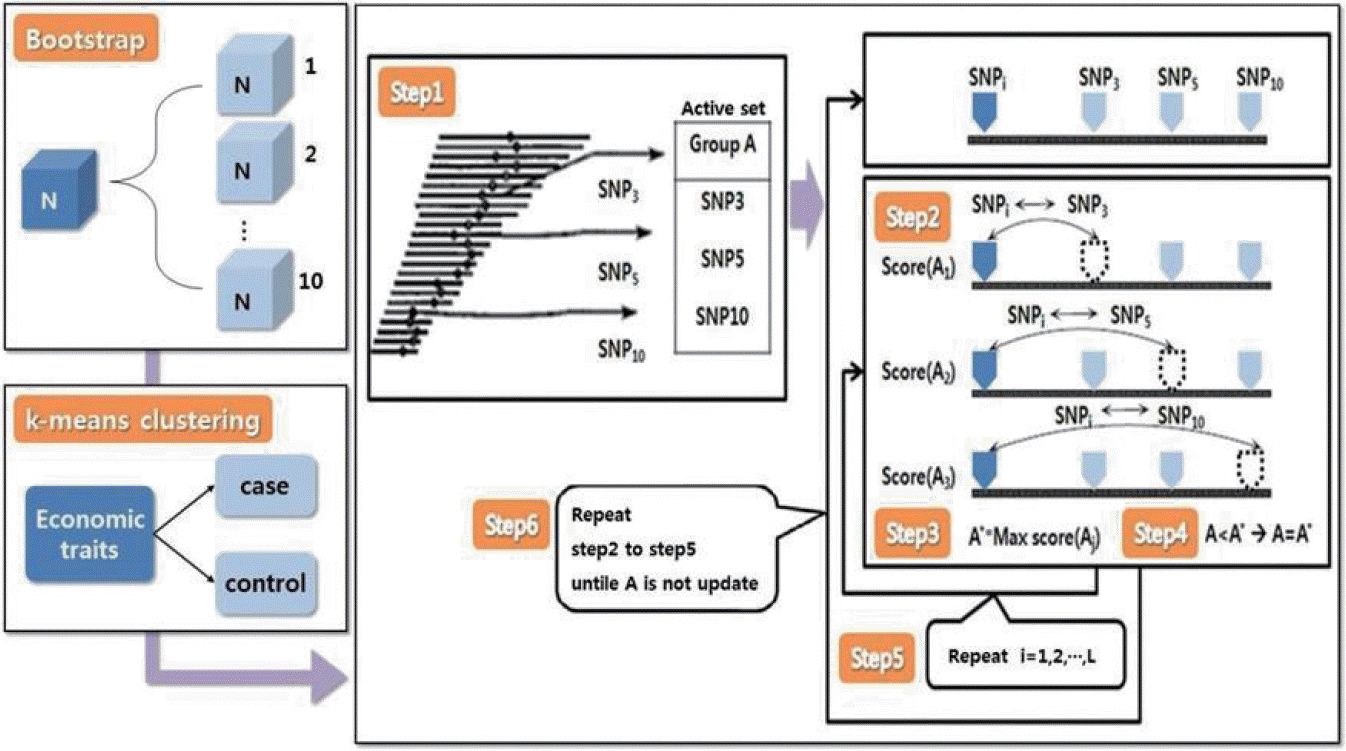
Step 1. Randomly select k groups from all SNP groups and assign group names (e.g., group A). Then set the rest of SNPs as SNPi.
Step 2. Exchange SNPi that do not belong to group A with group A elements on a one-by-one basis to calculate scores:
Step 3. Set the highest value from Step 2 as A*.
Step 4. If A* has a higher score than A, then replace A with A*.
Step 5. If the score of A* exceeds the threshold, then A* is classified as a significant group.
Step 6. For SNPi+1 that does not belong to group A, repeat Steps 2–5.
Step 7. If A* is not replaced with any other SNPi+1, then the process is stopped, and A* is determined as the final SNP combination set.
By repeating these steps, SNP combinations influencing economic traits were selected. The SNP Harvester method was applied only to binary data in the case-control study. Therefore, measures of economic traits were converted into binary values through k-means cluster analysis.
RESULTS AND DISCUSSION
In this study, the SNPs associated with the FAC and quality of Hanwoo were FABP4 (g.2634+1018 A>T, g.2988 A>G, g.3690 G>A, g.3710 G>C, g.3977-325 T>c, and g.4221 A>G) and SREBPs (g.3270+10274 C>T and g.13544 T>C). In addition, the four economic traits (C18:1, SFAs, MUFAs, and MS) that affect the evaluation criteria for Hanwoo, particularly the flavor and meatiness of beef, were considered. In addition, environmental factors such as age and the breeding farm were adjusted by linear regression analysis to evaluate only the genetic effect. Data were from 5,130 bootstrap samples based on the 513 steers in Oh et al [22]. The SNP Harvester method was used to find major gene combinations, and then a multifactor dimensionality reduction method was used to identify superior genotypes in gene combinations. Finally, the superior gene or SNP combination was identified by comparing individual and interaction effects.
Table 1 and 2 show the upper seven SNP combinations of two-way gene interactions related to the four economic traits. The value of x2 statistic used in SNP Harvester method, p-value and Cohen’s w were indicated. The effect size, w, was used by Cohen (1988) [30]. They set a small value of w at 0.1, a medium value at 0.3, and a large value at 0.5 [29]. Because most of the values shown in the Table 1, 2 appears larger than the medium value of 0.3, result can be seen to be effective. For the economic traits (C18:1, SFAs, and MUFAs) associated with fatty acids (Table 1), (g.3270+10274 C>T g.13544 T>C), (g.3270+10274 C>T, g.2634+1018 A>T), (g.2988 A>G, g.3977-325 T>C), and (g.3977-325 T>C, g.4221 A>G) were chosen as major gene combinations. For MS (Table 2), the upper three combinations (g.3690 G>A, g.3977-325 T>C), (g.3270+10274 C>T, g.3977-325 T>C), and (g.2634+1018 A>T, g.3690 G>A) were selected as major gene combinations.

Upper seven-SNP combinations based on two-way gene interactions for each fatty acid

Upper seven-SNP combinations based on two-way gene interactions for MS
Table 3 and 4 show the analysis results for differences between superior genotypes and others for selected major gene combinations using the multifactor dimensionality reduction method. For C18:1 and MUFAs, an increase in their content increases the quality of Hanwoo [8,10], whereas the opposite trend arises for SFAs [22]. In all gene combinations, differences between superior genotypes and others were significant. In addition, (g.2988 A>G, g.3977-325 T>C) and (g.3690 G>A, g.3977-325 T>C) were selected as the best combinations with the largest difference in traits associated with fatty acids (C18:1, SFAs, and MUFAs) and MS, respectively (p<0.0001).
Environmental factors were adjusted for in order to evaluate only the genetic effect. Table 5 shows the adjustment effect by comparing the accuracy before and after correction in two-way gene interactions. There were increases in accuracy in all gene combinations. In addition, (g.2988 A>G, g.3977-325 T>C) and (g.3690 G>A, g.3977-325 T>C), which were selected as the best combinations, showed the largest increases. After adjustment, training- and testing-balanced accuracy values for (g.2988 A>G, g.3977-325 T>C) were 0.9210 and 0.9163, respectively, for C18:1; 0.9329 and 0.9329, respectively, for SFAs; and 0.9314 and 0.9314, respectively, for MUFA. Further, training- and testing-balanced accuracy values for (g.3690 G>A, g.3977-325 T>C) were 0.9143 and 0.9143, respectively, for MS. Table 6 compares the individual and interaction effects of the best combination. For all economic traits, g.3977-325 T>C was the superior gene, and CC of g.3977-325 T>C was the superior genotype. For C18:1 and MUFAs, the g.3977-325 T>C and (g.2988 A>G, g.3977-325 T>C) combinations had the same effect.

A comparison of individual and two-SNP interaction effects on fatty acids and MS
In conclusions, g.3977-325 T>C and (g.2988 A>G, g.3977-325 T>C) combinations of FABP4 were the superior SNP, particularly combinations of the CC genotype of g.3977-325 T>C and the AACC, GACC, GGCC genotypes of (g.2988 A>G, g.3977-325 T>C). Consequently, future research should focus on this gene to produce high-quality beef that is low in SFAs and high in MUFAs and MS for the further development of the Hanwoo industry.
Notes
CONFLICT OF INTEREST
We certify that there is no conflict of interest with any financial organization regarding the material discussed in the manuscript.