Growth Performance, Carcass Characteristics and Plasma Mineral Chemistry as Affected by Dietary Chloride and Chloride Salts Fed to Broiler Chickens Reared under Phase Feeding System
Article information
Abstract
Requirements of dietary chloride (dCl) and chloride salts were determined by using 4×2 factorial arrangement under four phase feeding program. Four levels (0.31, 0.45, 0.59 and 0.73%) and two sources (NH4Cl and CaCl2) of the dCl were allocated to 1,472 chicks in eight dietary treatments in which each treatment was replicated four times with 46 birds per replicate. The four phase feeding program was comprised of four dietary phases: Prestarter (d 1 to 10), Starter (d 11 to 20), Grower (d 21 to 33) and Finisher (d 34 to 42); and diets were separately prepared for each phase. The cations, anions, pH, dissolved oxygen (DO), temperature, electrical conductivity (EC), total dissolved solids (TDS) and salinity were analyzed in drinking water and were not affected by dietary treatments. BW gain (BWG; p≤0.009) and feed:gain (FG; p≤0.03) were improved in CaCl2 supplemented diets during d 1 to 10. The maximum response of BWG and FG was observed at 0.38% and 0.42% dCl, respectively, for d 34 to 42. However, the level of dCl for BWG during d 21 to 33 (p≤0.04) and d 34 to 42 (p≤0.009) was optimized at 0.60% and 0.42%, respectively. The level of dCl for optimized feed intake (FI; p≤0.006), FG (p≤0.007) and litter moisture (LM; p≤0.001) was observed at 0.60%, 0.38% and 0.73%, respectively, for d 1 to 42. Water intake (DWI) was not affected by increasing dCl supplementation (p>0.05); however, the ratio between DWI and FI (DWI:FI) was found highest at 0.73% dCl during d 1 to 10 (p≤0.05) and d 21 to 33 (p≤0.009). Except for d 34 to 42 (p≤0.006), the increasing level of dCl did not result in a significant difference in mortality during any phase. Blood pH and glucose, and breast and thigh weights (percentage of dressed weight) were improved while dressing percentage (DP) and gastrointestinal health were exacerbated with NH4Cl as compared to CaCl2 supplemented diets (p≤0.001). Higher plasma Na+ and HCO3− and lower Cl− and Ca++ were observed in NH4Cl supplemented diets (p≤0.001). Increasing supplementation of dCl increased plasma Cl− (p≤0.04; quadratically) and linearly reduced plasma K+ (p≤0.001), Ca++ (p≤0.003), HCO3− (p≤0.001), and Na+ (p≤0.001; quadratically). Consequently, higher requirements of dietary chloride are suggested for feed intake; nevertheless, lower levels of dietary chloride are sufficient to support optimal BWG and FG with increasing age. The NH4Cl supplemented diets ameliorate breast and thigh meat yield along with overall energy balance (glucose).
INTRODUCTION
A dynamic relationship exists between cations and anions in order to maintain homeostasis in body fluids. Sodium (Na) and potassium (K) serves as major cations while chloride (Cl) and bicarbonate (HCO3) represents the anionic nature of these fluids. The cation-anion difference, known as dietary electrolyte balance (DEB; Mongin, 1980) can be described as the changes that occur in the body to achieve normal acid-base balance (ABB). The DEB has been considered as an important factor in broiler diets but becomes more critical when there is a high inclusion proportion of either cation or anion in the diet. Mushtaq et al. (2005 and 2007) keeping in view the concept of Mongin (1980), suggested that a level of 250 mEq/kg DEB is appropriate to balance cations and anions.
The supplementation of dCl has been investigated in poultry diets according to the age group, environmental conditions (temperature and relative humidity) and dietary composition (protein composition particularly). The National Research Council (NRC, 1994) recommended 0.20% and 0.15% dCl for starter (1 to 3 wk) and finisher (4 to 6 wk) phases of broiler growth, respectively. Under heat stress conditions, the concentration of Cl ions in the blood increases whereas concentration of Na and K decreases causing blood acidification that resulted in low blood electrolyte balance and increased blood Cl contents (Belay and Teeter, 1993; Borges, 1997; Ahmad et al., 2005). Borges et al. (2004a) found poor performance in birds with blood acidosis when the birds were fed high Cl (0.73%). Mongin and Sauveur (1974) reported a decline in weight gain with an increased level of dCl (i.e. 0.35, 1.01 and 1.41%) when Na and K were kept at 0.22% and 0.39%, respectively.
Various salts have been added to manipulate osmotic balance in broiler chickens and this change leads to changes in water consumption and excretion (Smith and Teeter, 1989; Borges et al., 2004a; b). When calcium chloride (CaCl2) is used to supplement dCl it appears that the Cl is exchanged with bicarbonate ions in the lower digestive tract to make the whole environment acidic with the excretion of calcium as calcium carbonate (Mongin, 1981). Ahmad et al. (2005) indicated better growth performance and lower pH with ammonium chloride (NH4Cl) supplementation in heat-distressed broilers. Most of the electrolytic studies were conducted under the high ambient temperature; nevertheless, the current improvement in housing facilities and genetics has necessitated the critical evaluation of the requirements for dietary chloride, DEB and chloride salts in the normal physiological state. Therefore, the present study was envisaged to evaluate the effect of supplementation of dietary chloride with the applicability of the DEB using two chloride salts sources on growth performance, carcass traits, water intake, litter condition and plasma mineral chemistry of fast-growing broiler strain under the phase feeding program.
MATERIALS AND METHODS
Birds’ husbandry
A total of fourteen hundred and seventy two 1-d old straight-run Hubbard broiler chicks (Hubbard×Hubbard) were allocated to 8 dietary treatments replicated 4 times in such a way that each replicate had forty six (46) birds. Each replicate pen was equipped with separate overhead, transparent and volume-graduated twenty liter (20 L) water bottles linked to a nipple drinker line. Water bottles were cleaned and filled with fresh water after measuring the water consumption on a daily basis. One flat bottom round feeder was provided for each experimental pen. Birds were housed in an environmentally controlled system where variation in temperature and relative humidity were recorded and maintained. Continuous light was provided 24 h for first 3 d and then 23 L:1 D light pattern was adopted for the rest of the experimental period. A 7.5 cm deep fresh sawdust was used as litter material over a concrete floor. For the first 3 days, house temperature was maintained at 32°C and thereafter reduced by 0.5°C per day until 24°C was attained at d 19. Birds were vaccinated against Newcastle Disease (ND) plus Infectious Bronchitis viruses at d 4, Infectious Bursal Disease virus at d 8 and again at d 14; Hydropericardium Syndrome virus at d 18 and ND-Lasota strain at d 22 following the locally designed vaccination schedule.
Dietary plan
A basal diet having dCl, Na and K as 0.17, 0.30 and 0.92%, respectively, corresponding to a DEB value of 320 mEq/kg was formulated (Table 1). Four levels of dCl (0.31, 0.45, 0.59 and 0.73%) were supplemented to the basal diet with either commercially available feed-grade calcium chloride (CaCl2; in powder form, containing 63.9% Cl) and ammonium chloride (NH4Cl; in granular form, containing 66.3% Cl). The four levels of dCl corresponded to DEB values of 280, 240, 200, and 160 mEq/kg, respectively.

A large batch of the basal diet was prepared for each phase and the experimental diets were prepared using the basal diet. The experimental period was divided into 4 phases i.e., pre-starter (d 1 to 10), starter (d 11 to 20), grower (d 21 to 33) and finisher (d 34 to 42) according to the recommendations of the Hubbard management guide (Table 1). The nutrient composition was not exactly followed as per the instructions of the Hubbard.
All the ingredients were assayed for their proximate composition (AOAC, 2005) prior to diet formulation and actual values were used in the formulation. The Na and K contents of each diet were analyzed by flame photometer (AOAC, 2005) and Cl by titration with AgNO3 (Lacroix et al., 1970). The Cl, Na and K content of the final diets were again verified prior to start of the experiment. The analyzed values of the experimental diets were found close to the expected values. The ME of each ingredient was calculated by the appropriate regression equation suggested by NRC (1994). Amino acid composition of each ingredient was calculated using AminoDat™ 3.0 Platinum (Degussa AG, Germany) based on the DM and CP contents of each ingredient. The amino acid composition of each diet met or exceeded the ideal amino acid ratio suggested by Han and Baker (1994). The experiment lasted for 42 d of age, offering mash diets throughout the experimental period.
Data collection
Live performance:
Data on feed intake (FI), BW gain (BWG) and feed-to-gain ratio (FG) was recorded for each phase. The feed was withheld for six hrs before weighing the birds at the end of each phase to ensure the emptying of the digestive tract of the bird. Water intake (DWI) was recorded on a daily basis and a ratio between DWI to FI (DWI:FI) was also calculated for each phase. Mortality was recorded on a daily basis and dead birds were weighed prior to removal to correct FG.
Litter moisture:
Litter was collected at the end of each phase to determine its moisture (LM). For this purpose, about 500 g litter sample was randomly collected from different locations in each replicate pen. Each sample was homogenized, a representative sample of 100 g was taken, and oven dried at 105°C for 24 h (AOAC, 2005) to determine moisture contents.
Water analysis:
Water was analyzed for its pH by pH metre (LT-Lutron pH-207 Taiwan), dissolved oxygen (DO) by DO metre (YSI 55 Incorporated, Yellow Springs, Ohio, 4387, USA). Moreover, temperature, electrical conductivity (EC), total dissolved solids (TDS) and salinity were recorded by the Combo meter (H M Digital, Inc. CA 90230). These observations were recorded twice (morning and noon) daily from different replicates.
Carcass, blood and plasma mineral responses:
At the end of 42 d, two birds were randomly selected from each replicate for carcass and plasma mineral responses. Blood was collected from the wing vein in an EDTA-coated vacutainer for immediate pH monitoring and for blood glucose. Blood plasma was separated by centrifugation of blood samples at 2,000×g for 15 min (Hayat et al., 1999; Ahmad et al., 2005) and was analyzed for mineral (Cl, Na, K, Ca, P, Mg, HCO3) contents. The same birds were further used for evaluation of carcass characteristics. Carcass responses were evaluated in terms of dressing (% of live weight), breast, thigh, intestine, proventriculus and gizzard weights (% of dressed weight).
Statistical analyses
The experiment was executed under completely randomized design with 4×2 factorial arrangement using 4 levels of dCl from 2 sources of salt. Each pen mean was an experimental unit. The data obtained at the end of the experiment were subjected to ANOVA technique using GLM of Minitab 15.1 (Minitab Inc., State College PA) where linear, quadratic and cubic terms were studied in the model using polynomial contrasts. The level of significance was 0.05 unless or otherwise stated.
RESULTS AND DISCUSSION
The drinking water was analyzed daily for its EC (1.06 to 1.39), TDS (1,010 to 1,234), pH (7.17 to 7.49), salinity (0.8 to 1.0), DO (3.41 to 5.10) and temperature (24.4 to 33.1), keeping in view the concept that these water characteristics and concentration of the minerals (cations plus anions) could maneuver the DEB of ingesta (Teeter and Smith, 1986; Borges et al., 2003a, b). The water pH values were found within the normal range (6.0 to 8.5) that has been considered as optimal for broiler performance (Socha et al., 2002; Borges et al., 2003a, b). Moreover, the water TDS levels between 1000–3000 ppm are considered satisfactory for broilers (Chiba, 2009).
The response of dCl on BWG in different phases is presented in Table 2. A highly significant rise in BWG (p≤0.009) was observed by replacing NH4Cl with CaCl2 during 1 to 10 d of age. During d 11 to 20, a linear drop in BWG was noticed with increasing supplementation of dCl (p≤0.05). Significant cubic response in BWG was observed during d 21 to 33 (p≤0.04) and d 34 to 42 (p≤0.009). When optimized, maximum BWG (i.e. 702 and 579) was observed in 0.60% (DEB = 197 mEq/kg) and 0.42% (DEB = 248 mEq/kg) dCl, associated with the regression equation for d 21 to 33 (BWG = 924.8–1,770 Cl+3,983 Cl2−2,755 Cl3) and d 34 to 42 (BWG = −2,106+16,130 Cl−31,222 Cl2+19,138 Cl3), respectively. It is clear from these results that with increasing age, the requirements for dCl decreased and DEB increased as the dCl was dropped from 0.73% (DEB = 160 mEq/kg; d 1 to 10) to 0.60% (DEB = 197 mEq/kg; d 21 to 33) and then to 0.42% (DEB = 248 mEq/kg; d 34 to 42). This drop in elemental requirements of the dCl is in accordance with the recommendations of NRC (1994) but the numerical value is quite high in our experiment. This might be due to the fact that the levels recommended by NRC (1994) were determined using Vantress×White Plymouth Rock (Oliveira et al., 2005) which had more potential for mass and skeletal development than the fast-growing strains of today’s broiler. Secondly, the dCl requirements have not been investigated using four phases of the broiler’s life before this experiment. Murakami et al. (1997) in recent studies supported the view that the requirements of Na and Cl are increasing. Further, the higher levels of these minerals did not appear to restrict growth effects in terms of body weight and feed efficiency. It can be suggested from these observations that the requirements for dCl are higher than the previously mentioned levels and the role of dietary salt is not as important for growth performance if it does not change the rest of the nutrient formulations.

Effect of dietary chloride and chloride salts on body weight gain of broilers during various phases of the experiment
Cubic (p≤0.006) responses were observed for FI for the period of d 1 to 42 (Table 3) however, maximum intake was achieved at 0.60% dCl (DEB = 197 mEq/kg) when optimized using regression equation (FI = 3,629–1,485 Cl+ 5,128 Cl2–4,297 Cl3). This means that the appetite of birds is satisfied up to a certain extent and after that, bird excretes excessive Cl in order to maintain acid-base balance. The direct impact of dCl on appetite has been noticed in previous experiments that suggest better FI by increasing Na and Cl (Branton, 1986; Murakami et al., 1997; El-Deek, et al., 2010). However, in the present study it is considered that the negative effects of higher dCl on growth parameters may be compensated by higher levels of both Na and K. Further, the cation-anion ratio has a strong impact on acid-base balance in broilers and a blood pH of 7.28 (Hurwitz et al., 1973) has been considered as ideal for maximum growth efficiency which is also evident from the present study. Additional research data are required to fully determine the precise requirement of individual electrolytes, especially chloride. However, as far as the phenomenon of feed intake is concerned, the manipulation of DEB may assist it as by increasing DEB the intake also increased and that might adjust blood electrolytes and the acid base balance. Neither sources nor interaction effects were observed to play a role in FI during the whole experimental period.

Effect of dietary chloride and chloride salts on feed intake of broilers during various phases of the experiment
Except during pre-starter phase, there was no difference in FG brought about by replacing NH4Cl with CaCl2 during the rest of the experimental period (p≤0.03; Table 4). The higher BWG in the CaCl2 supplemented diets showed that birds efficiently utilized feed during d 1 to 10. However, marked effects (cubic responses) of the dCl were noticed in FG during the finisher (p≤0.004) and overall (p≤0.007) periods. Upon optimization, improved FG was observed at 0.38% dCl (DEB = 259 mEq/kg) for finisher (2.15) and overall (1.92) periods, respectively. Increasing the level of dCl exacerbated lysine-arginine antagonism, which alters the acid base balance in birds (Kim et al., 1989) and that might be the reason for the poor FG at higher dCl supplementation in the present study. It is clear that the amount of the dCl is not directly responsible for changes in the growth efficiency of chicks and the requirements of the dCl are different for different growth parameters like BWG, FI and FG. It was evident that sources play a pivotal role in growth performance during initial days of life while after that birds become more sensitive to the amount of dCl. No other study has yet been carried out to evaluate dCl requirements during the phase feeding program consequently this finding indicates there is need to establish the biochemical processes involved in the different phases of broiler life.

Effect of dietary chloride and chloride salts on feed:gain of broilers during various phases of the experiment
Neither amount, source nor interaction responses were observed on DWI during the whole experimental period (p>0.05, Table 5). The ratio between DWI and FI (DWI:FI) was significantly affected by source and level interaction for d 1 to 10 (p≤0.05, Table 6). At the highest level of dCl (i.e. 0.73%), DWI:FI was found lowest in CaCl2 supplemented diets and highest in NH4Cl supplemented diets. The numerical value of DWI:FI ranged from 1.88 to 2.85 which is higher than the previously observed values (i.e. 1.6 to 2.0; Dozier et al., 2002; Singleton et al., 2004). the supplementation of minerals not only stimulated water intake, but also adjusted mineral losses. Further, the higher water intake might neutralize the high acidic condition caused by the increased supplementation of the dCl. A cubic response was observed in DWI:FI during d 21 to 33 (p≤0.009) and found to be highest at 0.73% dCl. It is clear that this ratio was affected by the numerical change in FI during this phase. Ahmad et al. (2005) and Mushtaq et al. (2005, 2007) observed no response of dCl on DWI:FI when comparing different levels, electrolytes or salts, respectively.

Effect of dietary chloride and chloride salts on water intake of broilers during various phases of the experiment

Effect of dietary chloride and chloride salts on water intake-to-feed intake ratio of broilers during various phases of the experiment
Increasing supplementation of dCl (p≤0.04) and replacing NH4Cl with CaCl2 (p≤0.001) aggravated the litter moisture condition during d 1 to 42 (Table 7). Likewise, Pesti et al. (1999), Murakami et al. (2003) and Mushtaq et al. (2005, 2007) supports our finding. It is anticipated that poultry litter wetness could be affected by dietary nutrients (mainly electrolytes), health of gastrointestinal tract (GIT), age of the birds, regional production difference and various environmental factors (Patterson et al., 1998). In the present study, the negative effects of higher dCl are mainly attributed to its corresponding cation i.e. Na, the imbalance between these two ions cause excretion of more water in the faeces due to an imbalance of extracellular fluid and abnormal kidney function (Freeman, 1983). It is known that aldosterone plays major role in the retention of Na, Cl and water by the kidneys. These factors, in combination, resulted in a profuse body fluid volume that stimulated the release of atrial natriuretic peptide and aggravated the loss of fluid in the urine.
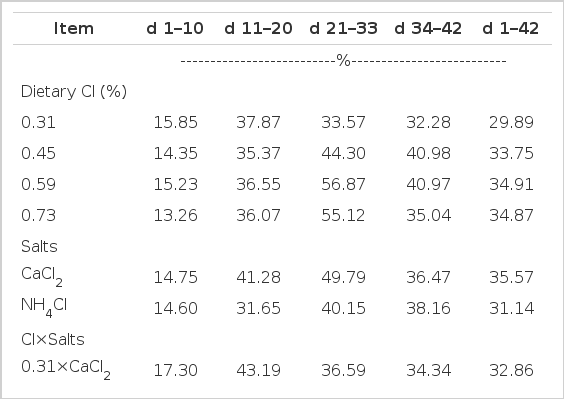
Effect of dietary chloride and chloride salts on litter moisture of broilers during various phases of the experiment
The highest mortality was observed in the highest level of dCl (0.73% or DEB = 160 mEq/kg) during d 34 to 42 (p≤0.003; Table 8). These results are in accordance with the findings of Ahmad et al. (2008) who observed high mortality in the diet having the lowest level of DEB (150 mEq/kg). This indicates that lighter birds can resist a higher dCl than heavier birds. Mortality remained unaffected in the rest of the experimental phases (p>0.05).
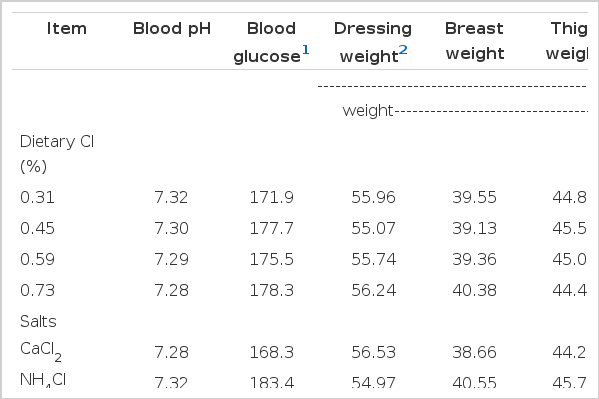
Effect of dietary chloride and chloride salts on blood and carcass responses of broilers at the end of the experiment
In the present study, the blood pH abated gradually with the increasing supplementation of dCl (p≤0.001; Table 9). An acidogenic role of dCl is well known as reported in a previous study by Murakami et al. (2001). However, a sharp reduction in pH value in CaCl2 supplemented diets might indicate the reduced absorption of Ca in the distal part of the intestine and more excretion as CaCO3, so Cl is retained to reduce blood pH (Mongin, 1981). It means that the role of dCl is more sensitive in the case of CaCl2 rather than in NH4Cl supplemented diets in order to maintain acid base balance. A profound decrease in blood glucose level was observed in CaCl2 supplemented diets compared to NH4Cl supplemented diets in the present study (p≤0.001). At the highest level of dCl (i.e. 0.73%), CaCl2 diets showed a lower glucose level while NH4Cl diets showed a higher glucose level (p≤0.04). Ahmad et al. (2005) also found reduced blood glucose in CaCl2 supplemented diets when compared with other ionic salts. A better glucose level overall depicts a better health status, however it is clear from the present study that the bird maintains its acid base balance at the cost of better health and growth.
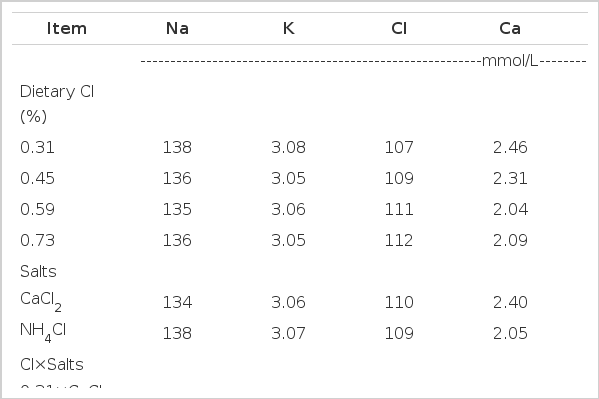
Effect of dietary chloride and chloride salts on plasma mineral chemistry of broilers at the end of the experiment
The CaCl2 supplemented diets proved to have a positive effect on dressing percentage (DP) when compared with NH4Cl supplemented diets (p≤0.02; Table 9). The CaCl2 supplemented diets showed highest DP (i.e. 57.62%) while NH4Cl showed lowest DP (i.e. 56.58%) at 0.31% dCl in the present study (p≤0.04). This increase in DP is evident from the higher intestinal weight, which is the indication of better gut health in CaCl2 supplemented diets in the present study. Moreover, the larger size of the proventriculus and gizzard (however numerically) may have improved the digestive capacity which resulted in higher DP in CaCl2 supplemented diets. Interestingly, the breast (p≤0.001) and thigh (p≤0.001) meat (% of dressed weight) was found higher when CaCl2 was replaced with NH4Cl in the experimental diets. Ahmad et al. (2005) also found higher breast meat yields in NH4Cl supplemented groups when comparing with other ionic salts. However, it is still unclear why DP has no relationship with breast and thigh meat yields. The data of body organs have been omitted as they are confusing the growth results.
The supplementation of dCl affected the plasma mineral chemistry of broilers as shown in Table 10. The higher plasma Na+ and HCO3− and lower Cl− and Ca++ were observed in NH4Cl supplemented diets (p≤0.001). The increase in HCO3− concentration is associated with a decrease in serum Cl− and an increase in the blood pH (Hurwitz et al., 1973; Ahmad and Sarwar, 2006). The utilization of Ca is exacerbated with metabolic acidosis, which is reflected from high urinary Ca in rats when diets were supplemented with NH4Cl (Bushinsky et al., 1982). A quadratic response in plasma Na+ (p≤0.001) and Cl− (p≤0.04) was noted with increasing level of dCl while plasma K+ (p≤0.001), Ca++ (p≤0.003), and HCO3− (p≤0.001) were linearly decreased with increasing supplementation of dCl in the present study. Plasma Cl− tended to be greater in birds receiving high levels of dCl and the electrolyte concentration in plasma is the reflection of the respective supplemental electrolyte in the diet (Kim et al., 1989; Ahmad et al., 2005).
CONCLUSION
It is inferred from the present study that a higher concentration of dietary chloride is required for feed intake; nevertheless, the lower levels are sufficient to sustain BWG and FG with increasing age. The NH4Cl supplemented diets ameliorated breast and thigh meat yields and the overall energy balance (glucose). It would be useful to test higher concentrations of dietary chloride with changing sodium and potassium ions within a constant electrolyte balance.
Acknowledgements
The present experiment was conducted and sponsored by the Research Model Broiler Unit, Poultry Research and Training Center, Department of Poultry Production, University of Veterinary and Animal Sciences, Ravi Campus, Lahore, Pakistan. The Higher Education Commission (HEC), Islamabad, Pakistan under the program ‘Indigenous PhD Fellowship Program (5000 Fellowships) Batch-V’ provided with the funding for chemical analyses of diet and blood. The contribution of Zulfiqar Ali (Ex-Nutritional Consultant, Big Feeds (Pvt.) Ltd., Lahore, Pakistan) in experimental diets preparation and analyses is highly appreciated. The authors also express gratitude to Rural Development Administration (RDA, Republic of Korea) for bearing the publication charges of this manuscript.