



INTRODUCTION
Methane is the primary greenhouse gas produced from livestock production, accounting for 37% of total anthropogenic CH4 emissions (FAO, 2006). Methane production is essential for effective degradation of organic matter, but also represents an energy loss of 2 to 12% gross energy intake (Johnson and Johnson, 1995). Consequently, strategies which can mitigate CH4 emissions, not only benefit the environment, but are of benefit to the animal as well.
Increasing interest in biodiesel, as a replacement for fossil fuels, has oilseeds, suitable for direct human consumption, being used for the production of renewable fuels. As such, the use of non-traditional grains and oilseeds, which do not compete directly with human consumption, are becoming attractive alternatives for the production of ethanol and biodiesel, respectively.
Biodiesel production yields two important by-products, oilseed press cakes and glycerine (Schröder and Südekum, 1999). The recent transition to the use of mechanical extraction produces oilseed press cakes with significantly higher oil contents leading to their use as energy feeds (Südekum, 2007). Similarly, glycerine, which forms the backbone of triglycerides and phospholipids in plants and animals, has been included in the diets of ruminant livestock for over 50 years at rates of up to 200 g/kg DM depending on purity (Johns, 1953; Schröder and Südekum, 1999).
Currently, increasing the lipid content of ruminant diets is considered the most effective way of consistently depressing ruminal methanogenesis (Grainger and Beauchemin, 2011). As oilseed press cakes are rich in fat, it follows that these biodiesel by-products could potentially exhibit a similar response, in terms of CH4 abatement, although there effects on digestion are largely unknown. Our objectives were therefore, to characterise the nutritive value of press cakes from moringa, castor, cotton, palm kernel, radish and sunflower and glycerine following oil extraction for biodiesel production and to examine their effects on in vitro ruminal fermentation, digestion kinetics and CH4 production.
MATERIALS AND METHODS
Substrates
Seven different oilseed press cakes commonly used for biodiesel production were tested in the in sacco study. These were: castor (Family Euphorbiaceae; Ricinus communis), cotton (Family Malvaceae; Gossypium hirsutum L), moringa (Family Moringaceae; Moringa oleifera Lam.), palm kernel (Family Euphorbiaceae; Elaeis guineensis, Jacq), radish (Family Brassicaceae; Raphanus sativus L. var. Oleiformes) and sunflower (Family Asteraceae; Helianthus annuus). Soybean meal [SBM, Family Fabaceae (alt. Leguminoseae; Glycine max)] and beard grass (Brachiaria brizantha) were used as a control.
The castor oilseed press cake was purchased from a Brazilian plant located in the municipality of Iraquara Ecodiesel, BA, and transported to Embrapa Semiarid, where it was detoxified using calcium hydroxide [Ca(OH)2] diluted in water at a ratio of 9 L of water/kg of Ca(OH)2 at a ratio of 60 g of Ca(OH)2/kg castor oilseed press cake. After treatment, the castor oilseed press cake was packed into a 200 L polyethylene drum for 24 h and allowed to dry in the sun for 12 h.
The remaining oilseed press cakes were obtained from Embrapa Dairy Cattle in Juiz de Fora (MG, Brazil). Glycerine was only tested in vitro and was obtained from ADM (Archer Daniels Midland) Company of Brazil Ltd (Rondonópolis, MT). Beard grass (Brachiaria brizantha; control) was harvested at 30 d of re-growth at the EMBRAPA Dairy Cattle Research Centre (Coronel Pacheco, MG, Brazil) located at 21° 35′ S, 43° 15′ W and 435 m above sea level. The DM and chemical composition of the oilseed press cake and beard grass samples were analysed (Table 1). The DM content of glycerine had been previously determined as 863.5 g/kg. Samples were dried and ground to 2 mm. Approximately 5 g DM of sample (Table 1) from each source was weighed into nylon bags (100×200 mm; pore size 50 μm) sealed and stored at room temperature.
In sacco study
Three rumen fistulated lactating Holstein-Zebu crossbred dairy cows were fed a diet consisting of beard grass forage supplemented with 2 kg of concentrate (22 g/kg CP and 12.6 g/kg NDF in DM) on a DM basis for the in sacco study. In sacco incubation measured the ruminal degradation of DM and crude protein (CP) of dietary treatments. For each cow, 24 weighted bags were placed at the bottom of the rumen, with three bags removed after 3, 6, 12, 24, 48 and 96 h of incubation. The 0 h bags were not placed in the rumen.
After removal from the rumen at the designated incubation times, nylon bags (including 0 h bags) containing diet residues were washed for 30 min on a cold rinse cycle (20°C) in a washing machine and then dried at 55°C for 48 h. Dried bags were weighed and residues were ground through a 1 mm screen for DM and CP analysis.
Digestion kinetics: Digestion curves for each data set were used to evaluate the in sacco degradation kinetics of dietary treatments in the rumen. Disappearance of DM and CP was analysed using a non-linear model as described by Lopez et al. (1999) to determine the fractional disappearance rate (k, h−1) and potential degradation (P) according to:
where A is the soluble fraction (proportion of each constituent, washed out of bags at t = 0 h), B is the insoluble degradable fraction, t is the time (h) and L is the lag phase (h). Effective degradability (ED) was calculated from the kinetic parameters obtained from exponential adjustment assuming a fractional passage rate (kp) of 0.02, 0.05 and 0.08 h−1:
The different turnover rates (kp) used here were based on maintenance feeding, medium and high production dairy cows (Chaves et al., 2006b), respectively.
In vitro incubations
Substrates: The substrates used for in vitro incubations were beard grass (Brachiaria brizantha; control) or beard grass supplemented with oilseed press cake or glycerine (0.864 w/v pure) in ratios of 1,000:0, 900:100, 800:200 and 600:400, for each treatment, respectively. Feed ingredients were dried at 55°C for 24 h and then ground to pass a 1 mm screen and mixed to obtain the 24 treatments. Each in vitro incubation was conducted as per Meale et al. (2012). Even at the 400 g/kg level of inclusion, the glycerine was fully absorbed onto the feed, leaving no free liquid. The entire incubation procedure was repeated twice (i.e., two incubation runs×three replicates per treatment, resulting in a total of six replicate vials per treatment).
Inoculum: Inoculum for the in vitro incubation was obtained from three ruminally fistulated cows grazing beard grass supplemented with 2 kg of concentrate (22 g/kg CP and 12.6 g/kg NDF in DM). Rumen fluid was collected 2 h before morning milking from 4 distinct sites in the rumen, filtered through 4 layers of cheesecloth, combined in equal portions from each animal and immediately transported in a pre-warmed Thermos® flask to the laboratory. Inoculum was prepared by mixing rumen fluid and a mineral buffer with 0.5 ml of cysteine sulphide solution (Vitti et al., 1999) in a ratio of 1:3. Inoculum (25 ml) was then transferred into pre-loaded, pre-warmed (39°C) vials under a stream of O2-free N gas. Vials were sealed and placed on an orbital shaker rack set at 120 oscillations per min in an incubator set at 39°C.
Determination of total gas, methane concentration and in vitro dry matter disappearance (IVDMD): Net gas production of each vial was measured at 6, 12, 24 and 48 h of incubation with a water displacement apparatus (Fedorak and Hrudey, 1983). At 6 h and 12 h prior to gas measurement, headspace gas was sampled from each vial with a 20 ml syringe and immediately transferred into a 5.9 ml evacuated Exetainer® (Labco Ltd., High Wycombe, Buckinghamshire, UK), which was then analysed for CH4 concentration by gas chromatography. Methane was expressed as mg of CH4/g of DM disappeared and total net gas production as ml/g of incubated DM. After gas was sampled for CH4 and total gas production was measured at 48 h of incubation, the fermentation vials were opened and the pH of the culture was measured using a pH meter (Orion Model 260A, Fisher Scientific, Toronto, ON, Canada). The ANKOM® bags with the residues were then removed from the bottles, rinsed thoroughly with distilled water and dried at 55°C for 48 h to constant weight to estimate IVDMD.
Determination of ammonia-N and VFA: A subsample (1.6 ml) of the culture media from each vial was transferred to 2 ml micro-centrifuge tubes and centrifuged at 14,000×g for 10 min at 4°C (Spectrafuse 16M, National Labnet Co., Edison, NJ, USA) to precipitate particulate matter and protein. The supernatant was transferred into 2 ml micro-centrifuge tubes and analysed for ammonia-N. In addition, a subsample (1.5 ml) of each vial was collected, acidified with 300 μl of metaphosphoric acid (0.25; w/v), and centrifuged as previously described for ammonia-N analysis. Supernatant was frozen at −20°C until analysed for VFA concentrations. The 0 h samples were also analysed for ammonia-N and VFA to calculate net ammonia-N and net total VFA production (Holtshausen et al., 2009).
Chemical analysis
Duplicate samples of each treatment and control diet were analysed for NDF and ADF using Van Soest et al. (1991) procedures modified for an ANKOM 200/220 Fiber Analyzer® (ANKOM Technol. Corp., Fairport, NY, USA), with heat-stable α-amylase. Concentrations of NDF and ADF were expressed inclusive of residual ash; NDF was corrected for CP and ash. Ash content was determined after 2 h of oxidation at 600°C in a muffle furnace (method 942.05; AOAC, 1990). Samples were re-ground using a ball grinder (Mixer Mill MM2000, Retsch, Haan, Germany) for determination of total N and the ADIN by combustion (method 990.03; AOAC, 1990). Crude protein was calculated as N×6.25. Ether extract (EE) content was ascertained by extraction with diethyl ether (method 920.39; AOAC, 1990) procedure modified for an ANKOM XT10 Extraction System® (ANKOM Technol. Corp., Fairport, NY, USA). Non-fibrous carbohydrate (NFC) was calculated as:
Statistical analyses
Univariate procedure in SAS was used to test for normal distribution of the data. Data were analysed as a randomized complete design using PROC MIXED (SAS, 2013). For the in vitro study, oilseed press cakes from biodiesel production treatment means for each level were compared against the control using the least squares mean linear hypothesis test (LSMEANS/DIFF) with the Dunnett adjustment with significance declared if p<0.05. For the in sacco study, data were analysed using a non-linear least-square procedure (PROC NLIN; SAS, 2103) to provide estimates for A, B and k calculated from DM and CP disappearance data. Data for all variables were analysed using the PROC MIXED procedure of SAS (2013) where treatments as fixed terms and cow and cow by treatment interaction as random effects. Significance was declared if p<0.05.
RESULTS
In sacco study
Dry matter digestion kinetics: The distribution of DM between soluble, degradable insoluble and undegradable residual fractions, disappearance rates (k, h−1) and effective degradability (EDDM; a prediction of degradation in vivo) of each supplement are summarised in Table 2. Soluble DM fraction was higher (p<0.05) for SBM, moringa and radish oilseed press cake than other treatments. Degradable insoluble DM was highest (p<0.05) for SBM, whereas the undegradable residual DM fraction was highest (p<0.01) for castor oilseed press cake when compared to other oilseed press cakes and beard grass (control). Kinetic data showed rapid degradation rates (k) of moringa oilseed press cake and sunflower and slow degradation of cotton oilseed press cake and beard grass. Moringa oilseed press cake had the highest (p<0.01) EDDM among treatments, closely followed by SBM and radish oilseed press cake, respectively (Table 2).
Crude protein digestion kinetics: Soluble CP fractions varied across all treatments (p<0.05). Radish and palm nut oilseed press cakes had the highest soluble fractions (p<0.05; Table 3). The insoluble degradable pool was highest for SBM and lowest for castor oilseed press cake (p<0.05). Conversely, castor oilseed press cakes and beard grass had the highest undegradable residual pool whereas, SBM and cotton oilseed press cake had the lowest (p<0.05). Sunflower oilseed press cake exhibited the greatest (p<0.05) fractional rate of CP disappearance when compared to all other treatments. Cotton and SBM had CP degradation rates (k) lower (average 0.04 h−1) than that of the beard grass control (p<0.05). Effective degradability of CP (EDCP) was greatest (p<0.01) for sunflower oilseed press cake, despite differences in distribution between pools, degradation rates and lag times (Table 3). Lag phases resembled patterns for DM, with longer (p<0.01) lag phase for SBM than the oilseed press cakes.
In vitro incubations
Replacing B. brizantha with castor oilseed press cake at 400 g/kg DM decreased gas production (ml/g DM) at 48 h (p<0.01; Table 4). However, no differences (p = 0.70) among treatments were observed in cumulative gas production, when expressed as ml/g digested DM. Glycerine produced the greatest amount of CH4 (mg/g digested DM; p<0.01) at both 6 and 12 h when included at 100 and 200 g/kg DM. Whereas, moringa oilseed press cakes, exhibited the lowest (p<0.01) CH4 production when compared to control, regardless of the inclusion level. Total VFA concentration did not differ (p≥0.30) between control and oilseed press cakes. However, there were differences (p = 0.04) among treatments, where 400 g/kg DM cotton oilseed press cake exhibited the highest total VFA concentration and 400 g/kg DM sunflower the lowest. The proportion of acetate in the total VFA increased (p<0.01) for castor oilseed press cake included at 400 g/kg DM yet decreased (p<0.01) for glycerine at 200 g/kg DM and 400 g/kg DM when compared to the control. Similarly, glycerine replacing B. brizantha at 200 g/kg DM and 400 g/kg DM produced higher (p<0.01) propionate levels compared to control. Castor oilseed press cake replacing grass at 400 g/kg DM decreased (p = 0.02) the proportion of butyrate in total VFA compared to control.
Replacing the grass control with glycerine and SBM, regardless of the proportion, improved IVDMD (p<0.01; g/kg) compared to the control. However, castor supplemented at 200 g/kg DM and 400 g/kg DM reduced (p<0.01) IVDMD. Ammonia-N (mg/dl) increased with the inclusion of all levels of radish oilseed press cake, 100 g/kg DM castor oilseed press cake and 400 g/kg DM of cotton, moringa and sunflower oilseed press cakes and SBM.
DISCUSSION
Dry matter in sacco characteristics
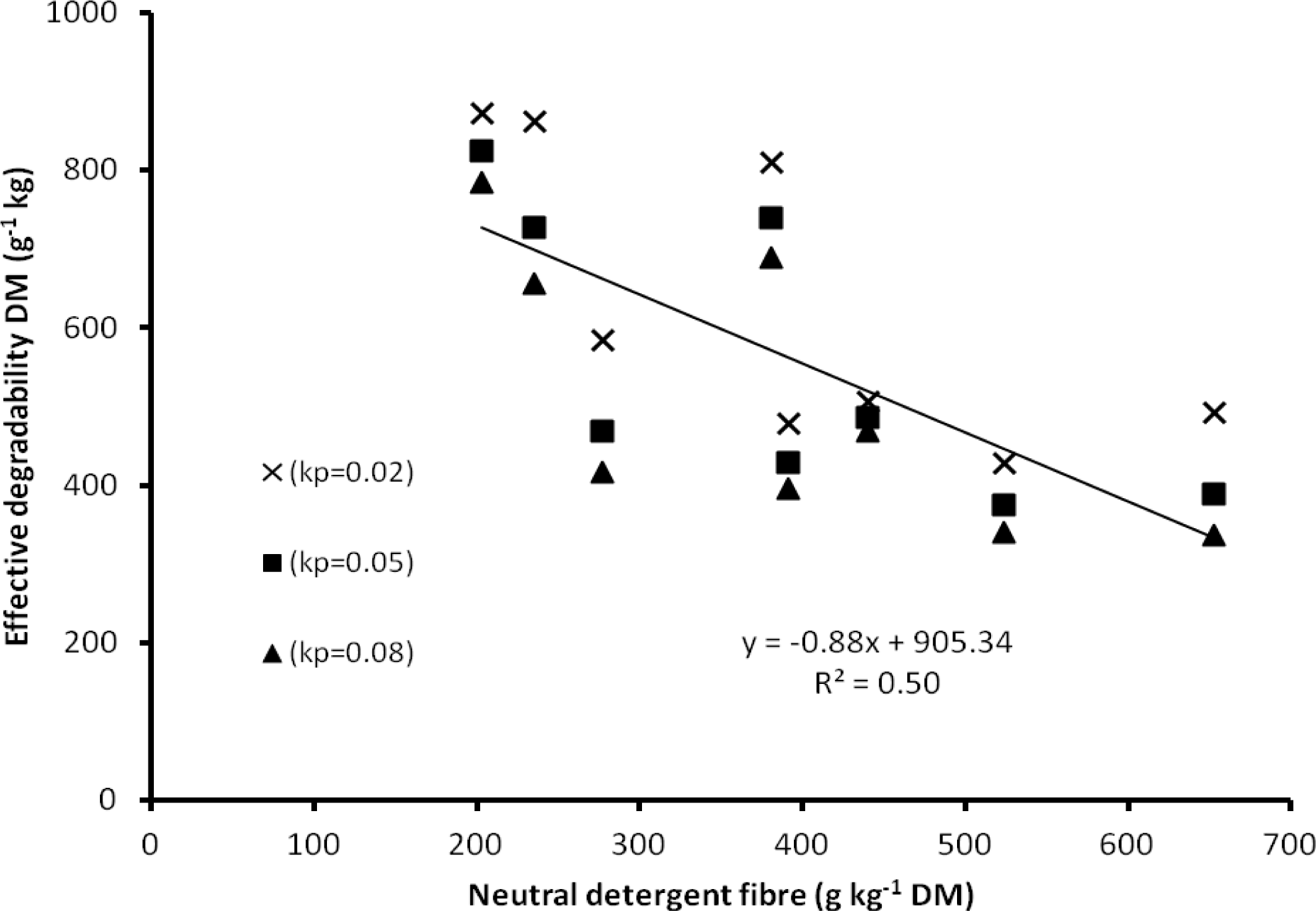
Lignification limits microbial access to structural polysaccharides in the cell wall resulting in slower digestion (Waghorn et al., 2003). Results of the current study support this, as there was a linear trend for supplements with higher concentrations of dietary NDF to exhibit a lower effective degradability of DM (EDDM; Figure 1). For example, the highest rapidly soluble fraction of DM was found in moringa oilseed press cake which also had the lowest concentrations of NDF and ADL (DM basis). Castor press oils seeds contained high levels of NDF and ADL, resulting in a low disappearance of the soluble fraction of DM, although the rate of DM disappearance (k, h−1) was moderate. High disappearance of the degradable fraction of DM in the rumen is nutritionally favourable as the supply of energy to rumen microbes increases (Chaves et al., 2006a).
A greater EDDM, such as that observed in moringa oilseed press cake, is preferential as the nutrients are more readily available and can therefore be more effectively utilised by the animal. In the current study, castor and palm kernel oilseed press cakes were the least degradable supplements. Both of these oilseed press cakes had high ADL and NDF concentrations (g/kg) indicating a lower nutritive value. This is in agreement with Gralak et al. (1997) which reported low rumen degradability of both DM and CP in palm seed meal. The presence of seed shells has also been reported to negatively affect degredation (Makkar et al., 2008) and whilst this may have had an effect in this study, the presence of seeds shells was not quantified.
Crude protein in sacco characteristics
The effective degradability of CP (EDCP) exhibited a similar trend to that of the EDDM whereby, increasing concentrations of NDF lead to lower EDCP (Figure 2) potentially having detrimental effects on microbial protein synthesis, as ruminally degradable protein supplies peptides, amino acids and NH3-N derived from proteolysis (Promkot et al., 2007). Moringa oilseed press cake exhibited the highest EDCP and a high rate of CP disappearance in the current study, most likely a result of its low NDF concentration and high CP content. The CP contents of moringa oilseed press cake (577.1±151.8 g/kg DM) were higher in the present study than previously reported for moringa oilseed press cakes (317 g/kg DM; Anwar and Rashid, 2007). The moringa used in the current study was completely devoid of bark, which may account for the differences observed. However, the EDCP of moringa oilseed press cake was comparable to that of sunflower, despite the considerably lower CP content of sunflower oilseed press cake. This may be a result of the higher soluble fraction of CP in sunflower oilseed press cake which suggests, that although the CP content is lower, it is still readily available.
Conversely, despite having a high CP content, cotton oilseed press cake had one of the lowest EDCP. Gohl (1998) reported cottonseed meal had relatively low rumen degradability, making it a good source of by-pass protein. Similarly, Sarwatt et al. (2004) reported a higher CP content in cotton oilseed press cake compared to moringa leaf meal (320 vs 274 g/kg DM), yet the soluble and degradable insoluble fractions of cotton oilseed press cake were significantly lower than for moringa leaf meal. The authors attributed this to a dilution of CP in the cotton oilseed press cake as a result of the inclusion of cotton seeds hulls during processing.
Variation in protein degradability may also reflect different proportions of structural and non-structural protein and carbohydrate fractions, which in turn affects their solubility and bio-availability. Castor oilseed press cake had a CP concentration similar to that of sunflower oilseed press cake, yet they exhibited the lowest and highest CP disappearance of all treatments, respectively. The EDDM of castor oilseed press cake was also low indicating that nutrients are not readily available from this biodiesel by-product. Conversely, Nagalakshmi et al. (2010) compared the inclusion of castor seed cake and groundnut cake (10 g/kg DM) in a complete lamb diet and concluded that castor seed cake did not affect nitrogen digestibility but increased the digestibility of crude fibre and NDF, when compared to groundnut cake.
In vitro incubations
Feed degradability is an essential component in determining the nutritive value of a feed. In the current study, the efficiency and extent of feed degradability was similar across all supplement types as reflected by the in vitro gas production. Increasing the level of supplementation of oilseed press cakes, SBM or glycerine to 400 g/kg did not affect gas production, when expressed as a function of digested DM, regardless of the supplement type. This suggests that the inclusion of these supplements in a forage based diet does not detrimentally affect digestion in a manner which is commonly associated with lipid supplementation, such as a reduction in in vitro bacterial growth and protozoal populations (Doreau and Chilliard, 1997), disturbances which can reduce gas production and DM digestion.
The supplementation of moringa and castor oilseed press cakes at 400 g/kg DM lowered gas production (mg/g DM). Interestingly, moringa oilseed press cake also had the lowest NDF and ADL contents of all treatments. These components, when present at high levels, are generally considered to impede microbial degradation of feeds (Johnson and Johnson, 1995). The presence of toxins, such as cationic polyelectrolyte proteins in moringa oilseed press cakes, which have been shown to exhibit antibacterial properties and bind strongly with rumen microbes, could potentially provide an explanation. At low levels, these proteins can protect feed proteins from degradation in the rumen, but, at high levels these proteins can inhibit rumen fermentation (Makkar et al., 2007) explaining the depression in gas production, although the levels of toxins present in each supplement were not measured in this study.
In vitro CH4 production is usually linked to digestibility, whereby highly digestible feeds are generally considered to produce more CH4 (Lee et al., 2011). This is supported by the results of the current study, where glycerine and SBM increased IVDMD (g/kg) above that of the control at all levels of inclusion. Correspondingly, glycerine produced the highest amount of CH4 (mg/g digested MD) at all levels of inclusion, with SBM showing an increase in CH4 at 400 g/kg DM, suggesting that CH4 production was indeed a reflection of diet digestibility. Although it is generally accepted that enteric fermentation of glycerine shifts the end products of fermentation towards the production of propionate, it is unlikely to form a hydrogen sink as occurs following carbohydrate fermentation and thus, a similar reduction in CH4 production was not observed. The inclusion of supplements which reduced the A:P ratio did not necessarily lower CH4 production. For example, inclusion of moringa oilseed press cake at 400 g/kg DM yielded the lowest amount of CH4, expressed in terms of digested DM yet, had an A:P ratio comparable to the control. Similarly, glycerine supplemented at this level had the lowest A:P ratio, yet exhibited the highest CH4 production at 12 h, suggesting an alternative method of CH4 suppression. Likewise, supplementing castor oilseed press cakes at 400 g/kg DM exhibited the highest A:P ratio yet no differences were observed in CH4 production (mg/g DM). These results are in agreement with Avila et al. (2011) which also observed a reduction in the A:P ratio without observing a reduction in in vitro CH4 (mg/g digested DM) when substituting increasing levels of glycerol for barley. Interestingly, castor oilseds press cakes had similar gas production to glycerine, indicating that a reduction in CH4 production was not a result of inhibited digestion. Although the castor oilseed press cakes used in the current study were detoxified, inclusion above 100 g/kg DM in a forage based diet reduced IVDMD with potentially detrimental effects on animal performance. Additionally, the use of castor oilseed press cake in ruminant diets is generally limited at 10 mg/kg, expressed in terms of castor-oil plant husks, due to the presence of the toxic glycoprotein, ricin (Madeira et al., 2011) indicating limited potential for inclusion of this supplement in ruminant diets.
Replacement of B. brizantha with oilseed press cakes, SBM or glycerine at up to 400 g/kg DM did not exhibit a similar effect to the supplementation of high levels of lipids in terms of reduced carbohydrate digestion. However, these supplements, with the exception of moringa oilseed press cakes at 400 g/kg DM, failed to inhibit CH4 production and are therefore considered to show limited CH4 abatement potential. Supplementation of Moringa oilseed press cake included at 400 g/kg DM was different in that, it lowered CH4 production, expressed as g/mg digested DM, without compromising nutrient degradation and consequently, exhibited the highest nutritive value of the supplements examined in this study.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print







