





INTRODUCTION
In regularly laying hens, there are three types of ovarian follicles differing in diameter, including slow-growing follicles (1- to 5-mm), prehierarchal follicles (6- to 8-mm diameter) and preovulatory hierarchy follicles (F6 to F1) [1]. During the process of follicle growth, after the recruitment of primary follicle, only some of prehierarchal follicles, also called small yellow (SY) follicles, will continue developing and finally ovulate. For egg laying, the F1 follicle will ovulate and subsequently one SY follicle with the highest expression of follicle stimulation hormone receptor (FSHR) will be selected to become hierarchal follicle [2]. This process is called follicle selection, which is an essential step impacting egg laying performance in chicken.
The SY follicles represent an important developmental stage in chickens, thus attract more attention. By Solexa sequencing, the expression patterns of the five miRNAs were analyzed in different developmental stages in chicken ovaries and in the follicles of various sizes including SY follicles [3]. Analysis on the transcriptomes of chicken SY follicles differing in FSHR mRNA expression indicated that Wnt signaling pathway was significantly enriched in the follicles with the greatest fold change in FSHR expression [4]. Global gene and protein expression in chicken SY follicles in response to acute heat stress was investigated and 176 genes and 93 distinct proteins with differential expressions were identified, among which the expression of heat shock proteins and peroxiredoxin family were upregulated [5]. Comparing gene expression between SY follicles in chickens and cattle at different follicular development stages revealed a role of estrogen receptor 2 in regulating cytochrome P450 family 19 subfamily A member 1 expression in the theca cells of chicken SY follicles [6]. During follicle selection from SY to F6 follicles, both the m6A methylation peaks and the m6A modified transcripts increased, which lead to the dynamic expression of many folliculogenesis relevant genes, including genes involved in Wnt signaling pathway [7]. Transcriptomic and proteomic comparison between chicken SY and F6 follicles revealed a role of very low density lipoprotein receptor in chicken follicle selection [8].
Some RNA molecules including miRNA [9] and lncRNA [10] play key roles in regulating the expression of protein coding genes. For chicken SY follicles differing in FSHR expression, we hypothesize that the differentially expressed genes (DEGs) between different follicles in the same individual might be regulated by miRNA and lncRNA. Therefore, in this study, by integrated transcriptomic analysis on the expression differences in the level of mRNA, miRNAs and lncRNAs in chicken SY follicles, we identified differentially expressed mRNA genes, miRNA and lncRNA genes. Gene interaction network analysis and subsequent functional in vitro test revealed that sosondowah ankyrin repeat domain family member A (SOWAHA) plays inhibitory role in chicken follicle selection.
MATERIALS AND METHODS
Animals and sample collections
Jining Bairi hens (a Shandong Indigenous chicken breed distributed mainly in Jining city) 35 to 40 weeks of age with regular laying for at least one month were used in this study. All sampled 60 Jining Bairi hens were killed by cervical dislocation immediately. Then, the ovaries including all sized follicles were carefully obtained from each chicken, prehierarchal follicles and hierarchal follicles were seperately collected to prepare granulosa and theca cells. The Institutional Animal Care and Use Ethics Committee of Shandong Agricultural University reviewed and approved all procedures described in this study (Permit Number: 2,007,005). This study was performed in accordance with the “Guidelines for Experimental Animals” of the Ministry of Science and Technology of China.
Library construction and sequencing
Total RNA was extracted from all of the SY follicles (6 to 8 mm) using TRIzol reagent (Invitrogen, Wilmington, DE, USA) and treated with DNase (TaKaRa, Dalian, China) to remove potential genomic DNA contamination, following the manufacturer’s instructions. The quantity and purity of the total RNA were evaluated. The RNA was then divided into two aliquots that were used for library construction of either small RNA or RNA. Fragmented RNA (the average length was approximately 200 bp) were subjected to first strand and second strand cDNA synthesis followed by adaptor ligation and enrichment with a low-cycle according to instructions of NEBNextUltra RNA Library Prep Kit for Illumina (NEB, Ipswich, MA, USA). The purified library products were evaluated using the Agilent 2200 TapeStation and Qubit2.0 (Life Technologies, Carlsbad, CA, USA). The libraries were paired-end sequenced (PE150, Sequencing reads were 150 bp) at Guangzhou RiboBio Co., Ltd. (Guangzhou, China) using IlluminaHiSeq 3000 platform.
Pre-processing of sequencing reads and quantification of gene expression level
Raw fastq sequences were treated with Trimmomatic tools (v 0.36) using the following options: TRAILING:20, MINLEN: 235 and CROP:235, to remove trailing sequences below a phred quality score of 20 and to achieve uniform sequence lengths for downstream clustering processes. Sequencing read quality was inspected using the FastQC software (v 0.11.6). Adapter removal and read trimming were performed using Trimmomatic (v 0.36). Sequencing reads were trimmed from the end (base quality less than Q20) and filtered by length (less than 25). Paired-end reads were aligned to the chicken reference genome GRCg6a with HISAT2. HTSeq v0.6.0 was used to count the reads numbers mapped to each gene. The whole sample expression levels were presented as RPKM (expected number of Reads Per Kilobase of transcript sequence per Million base pairs sequenced).
Gene ontology terms and KEGG pathway enrichment analysis of differential expressed genes
The statistically significant DE genes were obtained by an adjusted p-value threshold of <0.05 and |log2 (fold change)| >1 using the DEGseq software. Hierarchal clustering analysis was performed using the R language package gplots according to the RPKM values of differential genes in different groups, and colors represent different clustering information, such as the similar expression pattern in the same group, including similar functions or participating in the same biological process (BP). All differentially expressed mRNAs were selected for gene ontology (GO) and Kyoto encyclopedia of genes and genomes (KEGG) pathway analyses. GO was performed with KOBAS software (v 3.0). GO provides label classification of gene function and gene product attributes (http://www.geneontology.org). GO analysis covers three domains: cellular component (CC), molecular function (MF) and BP. The differentially expressed mRNAs and the enrichment of different pathways were mapped using the KEGG pathways with KOBAS3.0 software (http://www.genome.jp/kegg).
Co-expression network of differentially expressed lncRNAs and mRNAs
To investigate the potential functions of differentially expressed lncRNAs and the interactions between mRNAs and lncRNAs, we constructed lncRNA/mRNA transcripts co-expression network by calculating the Pearson correlation coefficient and p-value between multiple genes. In this study, the transcripts were filtered using a COR of >0.85 and a p-value of <0.05. Forty-four differentially expressed mRNA transcripts that were enriched in reproduction-related pathways and all differentially expressed lncRNAs were selected to construct a co-expression network that was illustrated using Cytoscape software (v 3.5.1).
Cell culture
Prehierarchal follicles were treated with 0.1% collagenase II (MP Biomedicals, Santa Ana, CA, USA) at 37°C for 8 min to disperse follicular granulosa cells and for an additional 30 min to disperse follicular theca cells. Theca cell and granulosa cell layers from each hierarchal follicle were collected and combined within their respective group and then dispersed for culture as previously described [4]. Hierarchal follicular granulosa cell layers were dispersed by treated with pancreatin (Gibco, Camarillo, CA, USA) for 15 min, while theca cell layers were dispersed with collagenase II for 30 min. The isolated theca and granulosa cells were planted in a 24-well culture plate containing 1 mL of M199 complete medium with high glucose (Gibco, USA) plus 10% fetal bovine serum (Biological Industries, Kibbutz Beit-Haemek, Israel).
Overexpression and knockdown assay
The entire coding region of chicken SOWAHA gene was amplified and polymerase PrimeSTAR (TaKaRa, China) was used to ensure high fidelity. The polymerase chain reaction (PCR) fragments were generated by double enzyme digestion and ligated with pcDNA3.1(+) expression vectors (Invitrogen, USA) by T4 DNA ligase, which were transformed into DH5α (TransGen Biotcch, Beijing, China) competent cells. After being confirmed by bidirectional sequencing and purified using an EndoFree Plasmid Purification Kit (Qiagen, Valencia, CA, USA), these plasmids were transfected into cells. Empty pcDNA3.1(+) vector was used as the control. For knockdown assay, siRNA was designed according to the chicken SOWAHA mRNA sequence (GenBank accession number XM_015294072). The negative control of the siRNA had the same composition with the siRNA sequence but had no homology with SOWAHA mRNA (Shanghai Gene-Pharma Co., Shanghai, China). The most effective siRNA was used to analyze the knockdown effect of siRNA on chicken SOWAHA gene.
Cell transfection and follicle stimulation hormone treatment
Granulosa cells were transfected with pcDNA3.1-SOWAHA overexpression plasmid or siRNA when grown to 80% confluency using NanoFectin Transfection Reagent (Shanghai ExCell Biology, Shanghai, China). As for FSH treatment, the cells were cultured with serum-free M199 medium in the absence or presence of different concentration of FSH (Sigma, St. Louis, MO, USA). Twenty-four hours after transfection and FSH treatment, the cells were lysed for RNA extraction.
Real-time quantitative polymerase chain reaction
The total RNA was extracted from all of the follicles using TRIzol reagent (Invitrogen, USA), and the total RNA from the cultured cells was isolated using a MicroElute Total RNA Kit (Omega, Norcross, GA, USA). The quality of the total RNA samples was tested by gel electrophoresis and spectrophotometry. The cDNA was synthesized using a Primescript RT reagent Kit with gDNA Eraser (TaKaRa, China), and the resultant cDNA was stored at −20°C for mRNA expression analysis. Real-Time quantitative PCR (qRT-PCR) was conducted on an MX3000p instrument (Stratagene, La Jolla, CA, USA) using the SYBR premix ExTaq (TaKaRa, China). Melting curves were used to confirm the specificity of each product, and the PCR efficiencies were determined by analysis of twofold serial dilutions of cDNA that were designed to detect all the signals in the spanning region. The efficiencies were nearly 100%, and therefore, the 2–ΔΔCT method for calculating the relative gene expression levels was used [11], and β-actin gene was used as the internal control. Primer sequences used for qRT-PCR are shown in Table 1.
Cell proliferation assay
The proliferation of granulosa cells was detected using an Enhanced Cell Counting Kit-8 Assay Kit (Beyotime, Beijing, China). Approximately 6×103 cells were seeded in every well of a 96-well plate. The cells were transfected with the pcDNA3.1-SOWAHA and empty pcDNA3.1 when the cells reached 60% confluence. At 0, 24, 48, and 72 h after transfection, 100 μL of medium with 10 μL of CCK8 was added to each well, and then the plates were incubated for a further 1 h at 38°C. The absorbance was evaluated using an ELx808 Absorbance Reader at 450 nm.
Statistical analysis
All data are presented as the mean±standard error of the mean. The differences between groups were determined by one-way analysis of variance followed by Duncan’s test using the SPSS software (SPSS Inc., Chicago, IL, USA). GO and KEGG analyses were assessed by Fisher’s t-test. For statistical analysis, p<0.05 was considered as significantly different.
RESULTS
Differentially expressed mRNA, lncRNA and miRNA genes between two small yellow follicles differing in FSHR expression in chickens
Previous studies suggest that among the eight to 10 SY follicles in laying hens, the one with highest FSHR expression will be selected to become hierarchichal follicle. Therefore, in this study, we compared the transcriptomes between two chicken SY follicles differing in FSHR expression. A total of 467 DEGs, including 331 upregulated and 136 downregulated genes were identified between two chicken SY follicles differing in FSHR expression at the significant criteria of (|log2 (fold change)| >1 and padj <0.001) (Figure 1A). KEGG analysis showed that these DEGs were mainly enriched in pathways of neuroactive ligand-receptor interaction and cell process-related phagocytosis related to environmental information processing (Figure 1B) and GO analysis showed that most of these DEGs were related to the function of cell composition in BPs (Figure 1C). Quantitative real-time PCR on 10 of these DEGs showed that their expression level was similar to the sequencing data, including SOWAHA, amphiregulin, BLK proto-oncogene, Src family tyrosine kinase, corin, serine peptidase, deoxyribonuclease 2 beta, centromere protein O, collagen type IV alpha 3 chain, family with sequence similarity 189 member A2, glucosamine (N-acetyl)-6-sulfatase, and relaxin 1, suggesting that the RNA-seq result was reliable (Figure 1D).
A total of 134 differentially expressed lncRNA genes, including 82 upregulated and 52 downregulated genes were identified between two chicken SY follicles differing in FSHR expression at the significant criteria of (|log2 (fold change)| >1 and padj <0.001) (Figure 2A). KEGG analysis showed that these differentially expressed lncRNA genes were mainly enriched in metabolic pathways and FoxO signaling pathways (Figure 2B) and GO analysis showed that most of these differentially expressed lncRNA genes were enriched in binding, cell and cell part, while more genes were significantly enriched in cell composition related to the function of cell composition in BPs (Figure 2C). Ten of these differentially expressed lncRNA genes (Figure 2D) were validated by qRT-PCR, which were similar to the sequencing data.
A total of 34 differentially expressed miRNA genes, including three upregulated and 31 downregulated genes were identified between two chicken SY follicles differing in FSHR expression at the significant criteria of (|log2 (fold change)| >1 and padj <0.001) (Figure 3A), and their target genes were predicted. One hundred and forty overlapped genes between the predicted target genes and DEGs were further analyzed by KEGG and GO enrichment analysis, showing that they were mainly enriched in advanced glycation end-recepor for advanced glycation end signaling pathway and neuroactive ligand-receptor interaction pathways (the pathways related to environmental information processing contain the most genes) (Figure 3B), and mostly were related to binding, single organism process, biological regulation, membrane and other significant functions (Figure 3C). Validation by qRT-PCR on 10 of these differentially expressed miRNA genes indicated that the results were consistent with the sequencing data (Figure 3D).
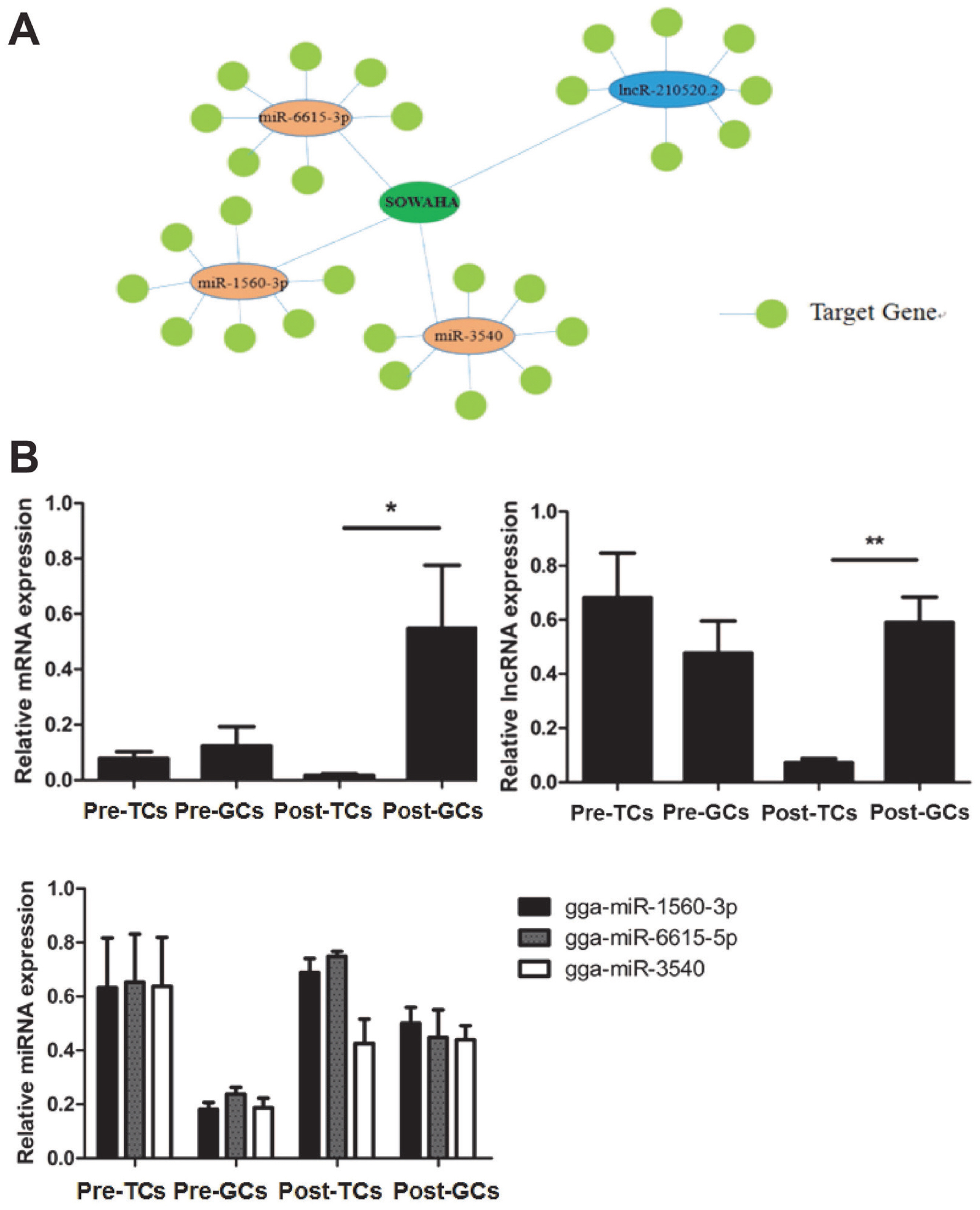
Integrated analysis on differentially expressed mRNA, lncRNA, and miRNA genes between two chicken SY follicles differing in FSHR expression
Integrated analysis indicated that SOWAHA is the common target gene of miRNA-6615-3p, miRNA-1560-3p, miRNA-3540 and lncRNA-210520.2 (Figure 4A). Sequencing results showed that, in follicles with higher FSHR expression level, the expression of SOWAHA and lncRNA-210520.2 was higher, while that of three miRNA genes was lower. While in follicles with lower FSHR expression level, the expression of three miRNA were higher, but that of SOWAHA and lncRNA-210520.2 was lower. The qRT-PCR analysis indicated that the expression of SOWAHA in hierarchal follicles was significantly higher in granulosa cells than in theca cells (Figure 4B). The expression of lncRNA-210520.2 in prehierarchal follicles was different from SOWAHA, while in hierarchal follicles, it exhibits similar trend as that of SOWAHA (Figure 4C). Changes in the expression of miRNA-1560-3p, miRNA-6615-3p, and miRNA-3540 in the granulosa cells and theca cells of hierarchal follicles were contrary to SOWAHA (Figure 4D).
Expression of SOWAHA gene in different follicles and changes after FSH treatment
The expression of SOWAHA was higher in SW follicles and SY follicles, markedly decreased in hierarchal follicles (F5, F3, and F1), and slightly increased in post-ovulation follicles (POFs) (Figure 5A). After FSH treatment, the mRNA expression of SOWAHA was significantly decreased (p<0.05) in granulosa cells of prehierarchal follicles, while it was increased (p<0.05) in hierarchal follicles granulosa cells after treatment with FSH at 5 ng/mL (Figure 5B).
Effect of SOWAHA on follicular development related genes in granulosa cells of prehierarchal and hierarchal follicles
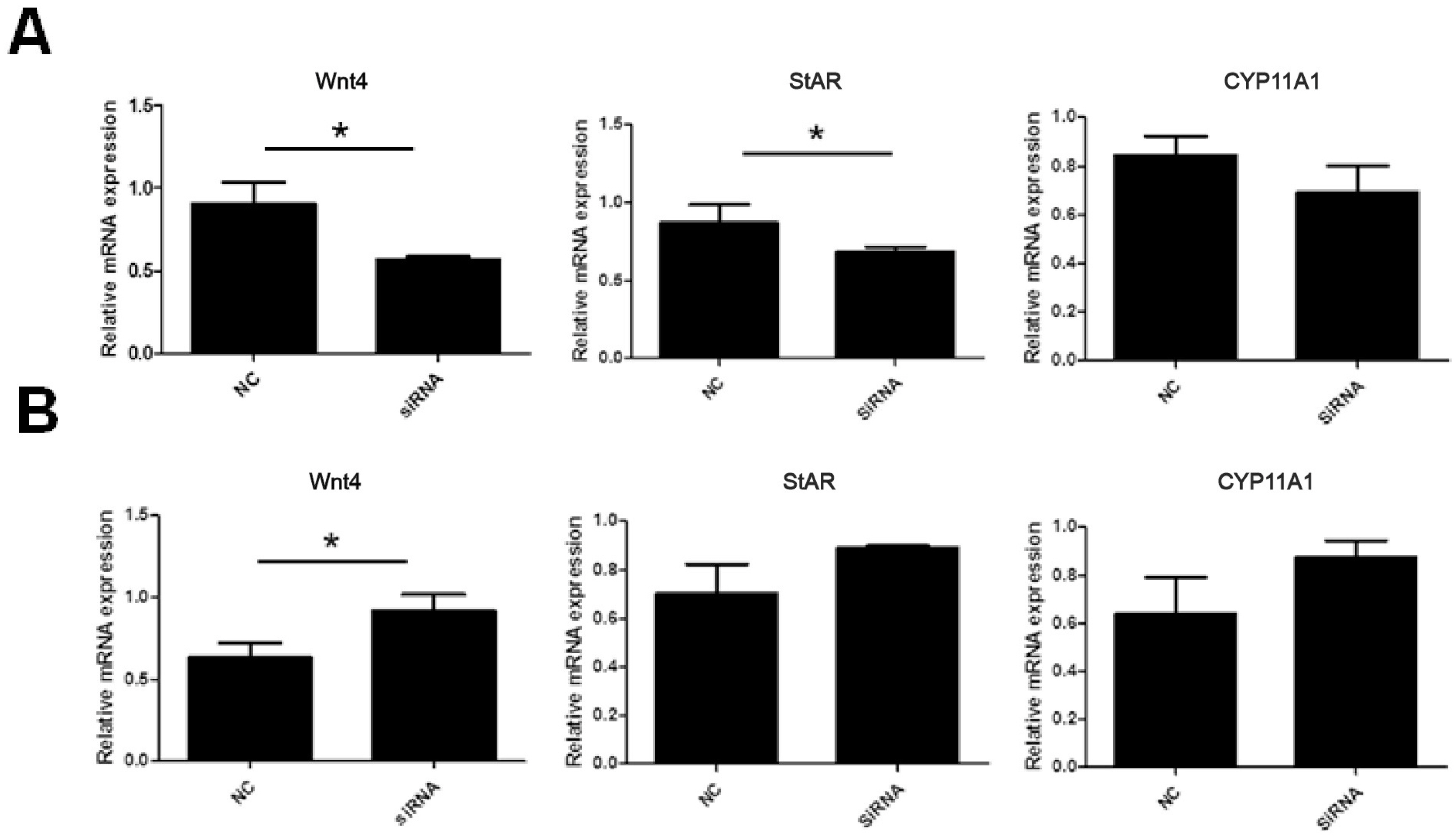
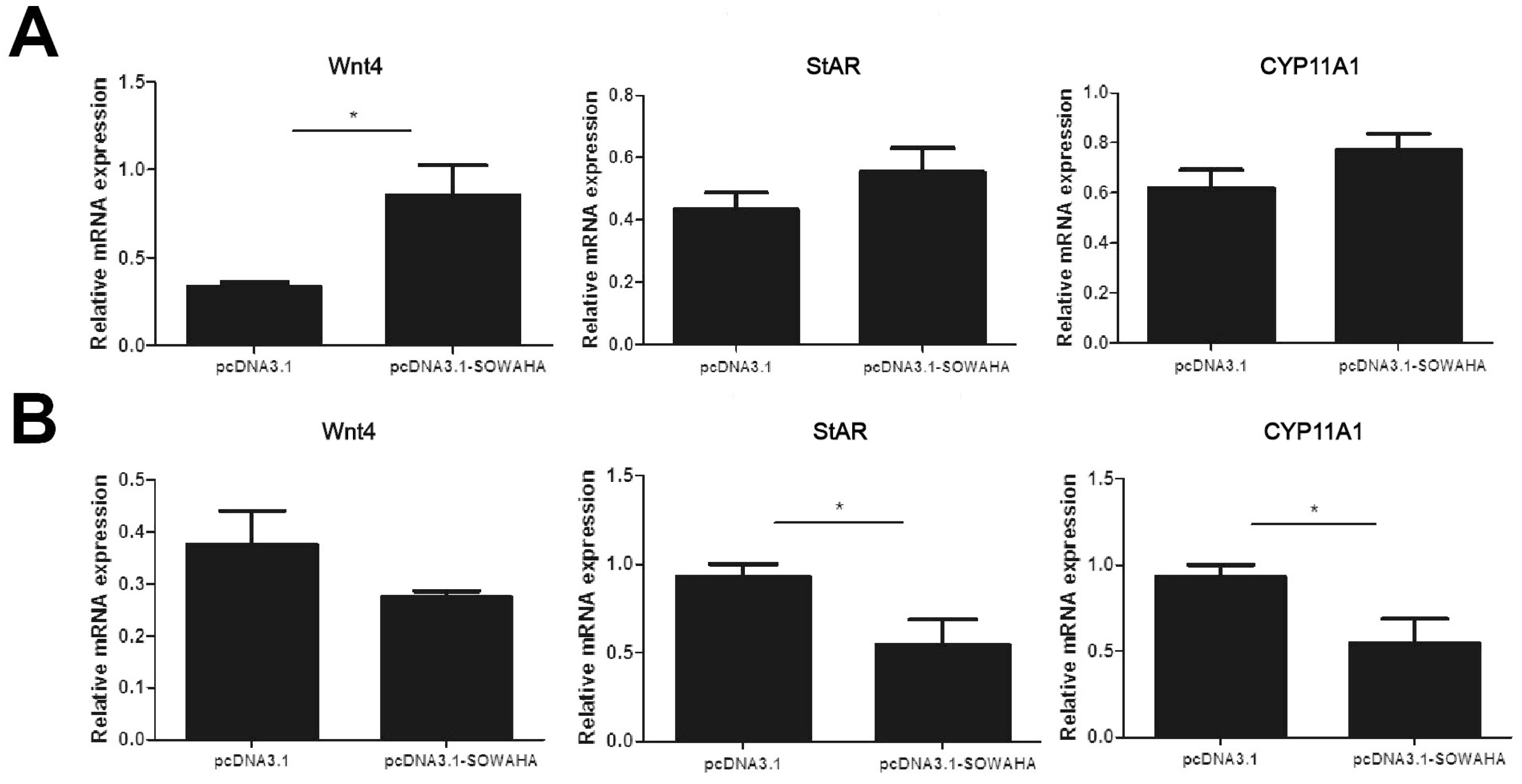
In the granulosa cells of chicken prehierarchal follicles, the expression of of Wnt family member 4 (Wnt4) and steroidogenic acute regulatory protein (StAR) mRNA was significantly decreased (p<0.05) when SOWAHA was knockdown by small interfering RNA (Figure 6A). However, after similar treatment, the expression of Wnt4 was significantly increased (p<0.05) in the granulosa cells of chicken hierarchal follicles (Figure 6B). After overexpression of chicken SOWAHA gene, the expression of Wnt4 was significantly increased (p<0.05) in prehierarchal follicles granulosa cells (Figure 7A), while the expression of StAR and cytochrome P450 family 11 subfamily A member 1 (CYP11A1) were significantly decreased (p<0.05) in hierarchal follicles granulosa cells (Figure 7B).
Effects of SOWAHA on the proliferation of chicken granulosa cells
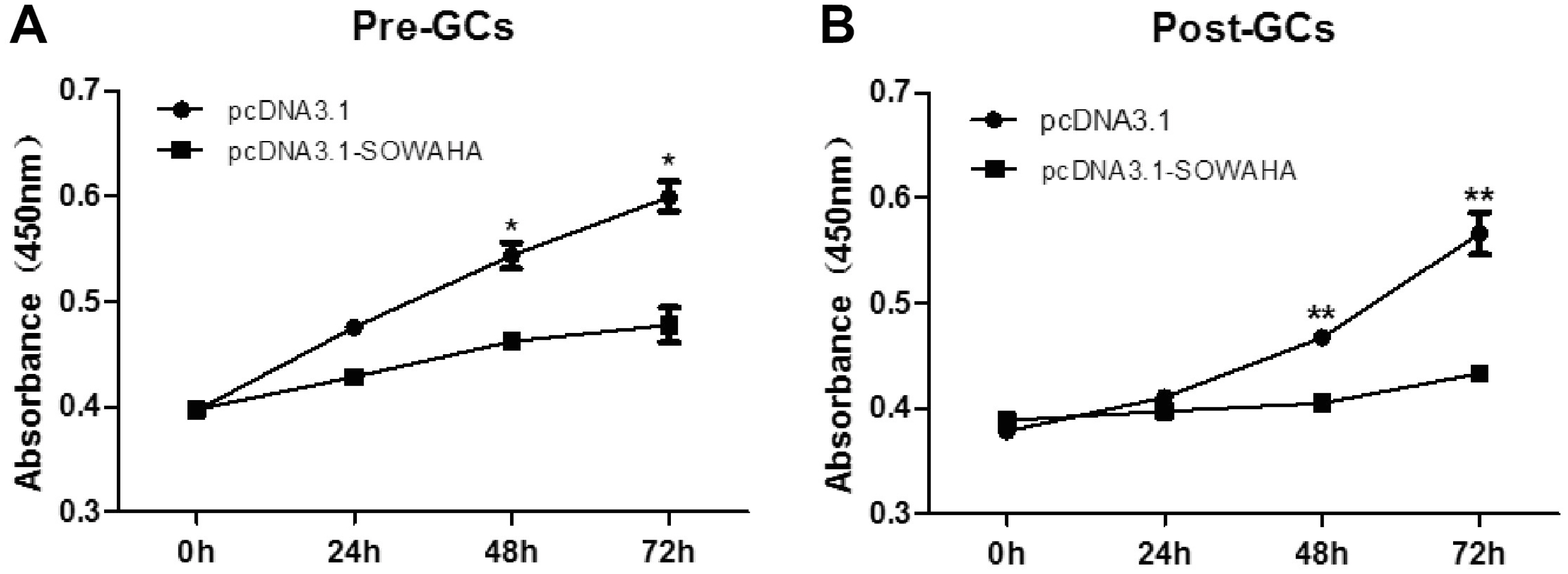
Granulosa cells were transfected with the overexpression vector pcDNA3.1-SOWAHA and empty pcDNA3.1 to examine the effect of SOWAHA on their proliferation. We found that overexpression of chicken SOWAHA gene significantly inhibited (p<0.05) the proliferation of granulosa cells from both prehierarchal (Figure 8A) and hierarchal follicles (Figure 8B).
DISCUSSION
Systemic transcriptomic analysis on expression differences between chicken SY follicles differing in FSHR expression is helpful for elucidating mechanisms underlying follicle selection in chicken. By previous study we uncovered the role of Wnt4 in chicken follicle selection [4]. In this study, for the first time, we performed an integrated transcriptomic analysis on these SY follicles, identified an interaction network of SOWAHA, miRNA-6615-3p, miRNA-1560-3p, miRNA-3540, and lncRNA-210520.2 and examined the effect of SOWAHA on Wnt4, StAR, and CYP11A1 mRNA expression, as well as on the proliferation of granulosa cells.
SOWAHA, also called ankyrin repeat domain-containing protein 43, is conserved in mammalian species including chimpanzee, Rhesus monkey, dog, cow, mouse, and rat, however, its homology with chicken SOWAHA is rather lower (33%). Genomic sequencing indicated that, despite 600 million years of independent evolution, Iroquois genes are linked to ankyrin-repeat-containing SOWAHA genes in nearly all studied bilaterians [12]. Three studies investigated the role of SOWAHA in mice and human. In HeLa-derived TZM-bl cells, knockdown of SOWAHA by siRNA inhibits HIV-1 replication [13]. During development, the expression of murine SOWAHA is observed in the postmitotic mantle zone during the specification and patterning of progenitor cells [14] and it is significantly downregulated by the exposure of nonsteroidal anti-inflammatory drugs to the canine melanoma cell line [15]. In this study, we found a dramatic decline of SOWAHA in hierarchal follicles and POFs than in prehierachical follicles and FSH treatment brought about different effect in granulosa cells of prehierarchal and hierarchal follicles. In SY follicles, the one with the highest FSHR expression would be stimulated by circulating FSH, causing a decline in SOWAHA due to the inhibitory role of FSH, and this SY follicle will be selected. In granulosa cells of hierarchal follicles, the effect of FSH on SOWAHA was not detected except at low concentration (5 ng/mL).
The expression of StAR and CYP11A1 is prerequisite for progesterone synthesis in the granulosa cells of chicken follicles, indicating the occurrence of granulosa cell differention, therefore, is the hallmark of follicle selection [16]. Wnt4 plays an important role in chicken follicle selection by stimulating granulosa cell proliferation and steroidogenesis [4]. Knockdown and overexpression of SOWAHA produced a different effect on the expression of Wnt4, StAR, and CYP11A1 in granulosa cells of prehierarchal and hierarchal follicle. In prehierarchal follicles, SOWAHA significantly stimulated the expression of Wnt4, but its effect on CYP11A1 was not significant. In hierarchal follicles, however, SOWAHA inhibits the expression of StAR and CYP11A1. Moreover, SOWAHA inhibits the proliferation of granulosa cells of both prehierarcha and hierarchal follicles. These results collectively suggest that, during follicle selection, with the increasing expression of FSHR, the function of SOWAHA would be inhibited and consequently, the expression of StAR and CYP11A1 and granulosa cell proliferation will increase. SOWAHA plays an inhibitory role in chicken follicle selection by affecting granulosa cell proliferation and differentiation.
Integrated analysis indicated that the expression of SOWAHA was regulated by three miRNAs, i.e. miRNA-6615-3p, miRNA-1560-3p, miRNA-3540, as well as by one lncRNA (lncRNA-210520.2) (Figure 4). The role of miRNA-6615-3p, miRNA-1560-3p, miRNA-3540, and lncRNA-210520.2 in ovarian follicles was not reported. During follicle selection in chicken, how SOWAHA is regulated by miRNA-6615-3p, miRNA-1560-3p, miRNA-3540, and lncRNA-210520.2 needs to be investigated.
In conclusion, in this study, by integrated analysis on chicken SY follicle transcriptomes, we identified SOWAHA as a network gene differentially expressed between SY follicles with marked FSHR expression. Further in vitro analysis indicated that SOWAHA was affected by FSH in granulosa cells of ovarian follicles. SOWAHA affected the expression of genes involved in chicken follicle selection and inhibited the proliferation of granulosa cells, suggesting an inhibitory role in chicken follicle selection.




 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print







